Loïc Legris , Olivier Detante et Nathalie Nasr
Le cerveau est un organe richement vascularisé. Le débit sanguin cérébral (DSC) correspond à environ 15 % du débit sanguin total, soit 700–750 mL/min. Le DSC dépend de la pression de perfusion cérébrale (PPC), elle-même dépendante de la pression artérielle (pression artérielle moyenne, PAM), et des résistances vasculaires cérébrales (RVC) en lien notamment avec le calibre des artères et artérioles. Ainsi, le DSC peut être calculé de la manière suivante :
DSC = PPC/RVC
Le métabolisme cérébral nécessite un apport sanguin constant en oxygène et en glucose. Les capacités de stockage énergétique sont pratiquement inexistantes, ce qui rend le cerveau vulnérable.
La vascularisation artérielle du cerveau est relativement variable et propre à chacun. Elle repose essentiellement sur des voies d'apport et des cercles anastomotiques existant à plusieurs niveaux, créant des réseaux et des suppléances, fonctionnellement très importants en particulier lorsqu'une voie d'apport est interrompue.
De nombreuses pathologies peuvent mettre à mal ce système artériel. En fonction de la sévérité et de la vitesse d'installation de ces atteintes, les systèmes de compensation sont dépassés et des lésions cérébrales ischémiques peuvent devenir irréversibles (infarctus cérébral ou AVC ischémique constitué). Les AVC sont ainsi la deuxième cause de mortalité dans le monde (6 millions de décès/an) et la première cause de handicap acquis de l'adulte.
La régulation cérébrovasculaire joue un rôle majeur dans la préservation du DSC. Le descriptif de la vascularisation cérébrale artérielle encéphalique est détaillé dans les chapitres 1 et 22, nous décrivons ici les mécanismes de régulation de la circulation cérébrale en physiologie et dans les situations pathologiques.
Vascularisation artérielle encéphalique
Le système artériel cérébral est issu de l'aorte et des différents troncs supra-aortiques (figure 27.1) d'où émergent quatre piliers : deux artères carotides (communes, puis internes) formant le réseau antérieur, et deux artères vertébrales qui s'unissent en une artère (ou tronc) basilaire unique pour constituer le réseau postérieur vertébrobasilaire. Ces réseaux s'anastomosent au niveau intracrânien via le cercle artériel de la base du cerveau (polygone de Willis), même si le caractère complet de ce « cercle » n'est pas la règle.
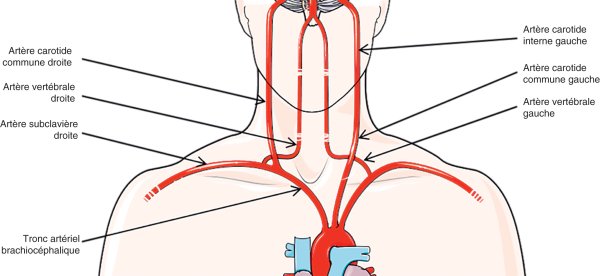
Figure 27.1
Troncs artériels supra-aortiques permettant la vascularisation de l'encéphale.
© D'après Servier Medical Art (2005).
Les artères carotides internes donnent naissance, en intracrânien, aux artères cérébrales antérieures et moyennes (ou artères sylviennes selon l'ancienne nomenclature), qui vascularisent la majeure partie de l'encéphale.
L'artère basilaire, quant à elle, se sépare en deux artères principales, les artères cérébrales postérieures. Elle donne aussi de nombreuses petites artères dites « perforantes » qui permettent la vascularisation du tronc cérébral et de territoires encéphaliques profonds.
Le cercle anastomotique artériel de la base du cerveau (figure 27.2) est variable selon les individus. Il permet des anastomoses antéropostérieures (artères communicantes postérieures) et entre les deux hémisphères (communicante antérieure reliant les deux artères cérébrales antérieures). Les variantes du cercle artériel de la base du cerveau sont fréquentes et portent majoritairement sur sa partie postérieure. Il peut s'agir par exemple d'une hypoplasie ou absence de la communicante postérieure ou bien d'une hypoplasie ou absence du 1er segment de la cérébrale postérieure avant la naissance de la communicante postérieure (dans ce cas, l'artère cérébrale postérieure naît de la carotide interne). Les variantes qui portent sur la partie antérieure du cercle artériel de la base du cerveau comme l'hypoplasie ou absence du 1er segment de l'artère cérébrale antérieure (dans ce cas, les deux cérébrales antérieures naissent de la même carotide interne), ou l'absence de la communicante antérieure ou le dédoublement de la communicante antérieure, sont moins fréquentes.
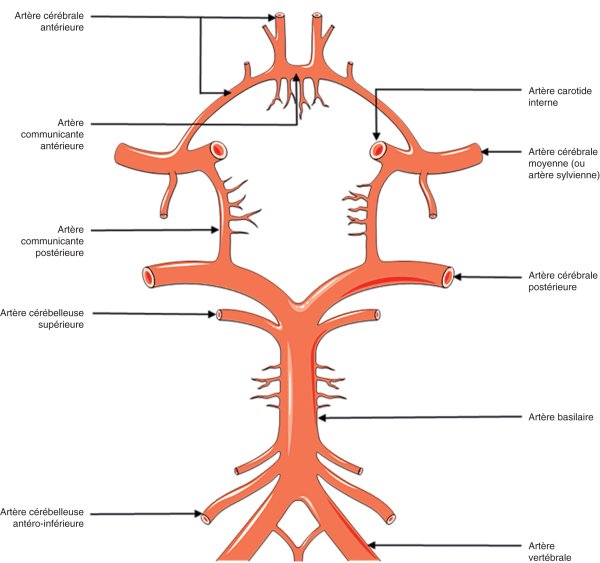
Figure 27.2
Cercle anastomotique du cercle artériel de la base du cerveau (polygone de Willis).
D'après Servier Medical Art (2005).
Deux autres types d'anastomoses existent : les anastomoses entre la carotide externe et la carotide interne via l'artère ophtalmique dont le flux s'inverse dans ce cas (en cas d'occlusion ou de sténose très serrée de la carotide interne bulbaire par exemple), les artères leptoméningées (ou piales) à l'origine d'anastomoses entre les territoires les plus distaux (par exemple le territoire de l'ACM [artère cérébrale moyenne] et celui de l'ACP [artère cérébrale postérieure] du même côté).
Régulation de la circulation artérielle cérébrale
Réseaux anastomotiques
Nous avons vu que les artères encéphaliques possédaient un certain nombre de communications entre elles, antéropostérieures, droites – gauches mais aussi avec le système carotidien externe.
Ces anastomoses permettent une protection de l'encéphale, pouvant s'ajouter à la redistribution cérébrale de secteurs vasculaires systémiques. Ainsi, lors de l'occlusion d'une artère, les vaisseaux perméables peuvent prendre le relais et permettre une compensation (souvent incomplète) du défaut d'oxygénation cérébrale.
Cependant, l'efficacité de ces réseaux est extrêmement variable selon les individus et les facteurs de risque neurovasculaire. Ces derniers altèrent au long cours les petites artères (par athérosclérose ou lipohyalinose), entraînant des lésions d'ischémie chronique, notamment de la substance blanche (leucoaraïose par maladie des petites artères) qui interfèrent avec la vasoréactivité et la récupération post-AVC.
Autorégulation artérielle
L'autorégulation peut se définir comme la propriété qu'ont les vaisseaux du cerveau de modifier activement leur diamètre afin d'adapter la RVC aux variations de la PPC, dans l'objectif de maintenir un DSC constant. L'autorégulation permet un plateau à 50 mL/100 g/min, via la mise en jeu de la réserve vasculaire composée principalement des réseaux artériolaires et capillaires (figure 27.3).
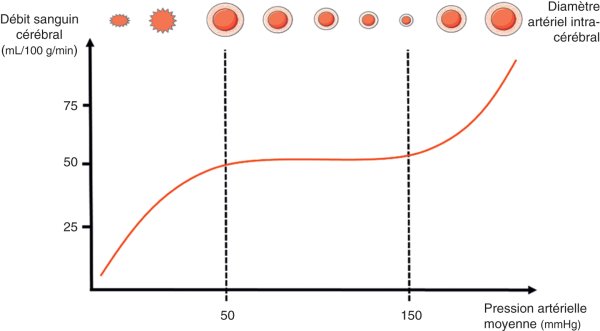
Figure 27.3
Courbe d'autorégulation cérébrale.
Mise en jeu des mécanismes de vasoréactivité permettant un plateau de débit sanguin cérébral autour de 50 mL/100 g/min.
© Loïc Legris.
Il s'agit ainsi d'un mécanisme physiologique de protection, tendant à limiter l'installation de l'ischémie en cas de modification importante de la PPC. Ce mécanisme a des limites inférieure (PAM < 60 mmHg) et supérieure (PAM > 150 mmHg), au-delà desquelles on constate une perte de l'autorégulation, le DSC suivant alors passivement les variations de PPC.
L'instauration de ces processus est très rapide (quelques secondes), et concerne à la fois les grosses et les petites artères. L'ajustement du calibre des vaisseaux à la pression de perfusion est lié aux propriétés des fibres musculaires lisses des parois.
Le muscle lisse de la paroi vasculaire réagit à l'étirement par une contraction (effet Bayliss), et inversement. Cet effet concerne principalement les grosses artères d'apport, ainsi que les artères corticales principales.
Facteurs de régulation
Le dioxyde de carbone (CO2) possède un rôle majeur de régulation du diamètre artériel. Ainsi, l'hypercapnie (augmentation de la PaCO2) entraîne un relâchement des fibres musculaires pariétales, responsable d'une vasodilatation et d'une augmentation du DSC. En cas d'hypercapnie, les vaisseaux sont donc dilatés, et perdent la capacité de se distendre davantage, modifiant la courbe d'autorégulation en la décalant vers la gauche voire à l'extrême en induisant une perte de l'autorégulation. À l'inverse, l'hypocapnie (diminution de la PaCO2) induit une vasoconstriction.
L'oxygène (O2) a aussi un impact important sur les modulations artérielles, notamment en cas d'hypoxie (diminution de la PaO2). Ainsi, lorsqu'elle est suffisamment sévère, cette dernière entraîne une vasodilatation avec augmentation du DSC. La dilatation est rapide (quelques minutes) et permet de maintenir constante l'extraction cérébrale d'oxygène.
Les facteurs humoraux sont moins présents (voire quasiment absents) en intracérébral du fait de la barrière hématoencéphalique.
La circulation cérébrale est aussi caractérisée par un couplage débit-métabolisme (neurovasculaire). Ainsi, l'activation d'une zone cérébrale induit une augmentation du débit sanguin, impliquant des mécanismes métaboliques complexes locaux. Ce phénomène explique que le DSC de la substance grise se situe autour de 80 ml/100 g/min, alors que celui de la substance blanche est plus faible, d'environ 20 ml/100 g/min.
Le potassium (K+), puissant vasodilatateur artériolaire, est l'un des principaux agents de ce couplage. Le K+ intracellulaire est libéré en grade quantité en extracellulaire lors de l'activation neuronale, induisant par une cascade une nouvelle libération aux pieds des astrocytes, au contact des microvaisseaux cérébraux.
Le monoxyde d'azote (NO) est lui aussi fortement impliqué dans le couplage débit-métabolisme. Radical libre gazeux synthétisé par des neurones actifs, il possède une action vasodilatatrice via le relâchement des fibres musculaires lisses.
Physiopathologie de l'ischémie cérébrale : de la pénombre à l'infarctus cérébral
Principales causes d'ischémie cérébrale
L'ischémie cérébrale est le plus fréquemment due à un embole d'origine artérielle ou cardiaque.
Au niveau artériel, il peut s'agir d'un agrégat plaquettaire (thrombus blanc) ou d'un thrombus fibrinocruorique sur une plaque d'athérome instable, voire d'emboles calciques ou de cholestérol. Par ailleurs, une occlusion localisée d'une petite artère perforante sur une plaque d'athérome intracrânien ou par lipohyalinose peut entraîner un infarctus profond de petite taille (lacune).
Au niveau cardiaque, il peut s'agir d'un thrombus fibrinocruorique endocavitaire (sur fibrillation atriale, valvulopathie, infarctus du myocarde), d'un embole paradoxal (d'origine veineuse) via un foramen ovale perméable (communication interatriale persistante après la naissance), voire plus rarement d'emboles calcique, septique (endocardite infectieuse) ou tissulaire (lésion tumorale).
D'autres causes plus rares peuvent être retrouvées : vascularite, vasospasme, hyperviscosité, thrombophilie, polyglobulie, etc.
Mécanismes physiopathologiques
D'un point de vue physiopathologique, l'ischémie cérébrale est définie comme une diminution de la PPC en dessous du seuil d'autorégulation du DSC. En dessous de 20 mL/100 g/min, on constate une insuffisance d'apport en O2 et en énergie. L'ischémie cérébrale est un phénomène évolutif dans le temps et l'espace, pouvant conduire très rapidement à des lésions irréversibles d'infarctus.
Dans un premier temps, la diminution du DSC entraîne une hypoperfusion avec maintien de la consommation en O2 grâce à l'augmentation de la fraction d'extraction en O2.
Par la suite, les mécanismes de compensation deviennent insuffisants, et la baisse du DSC est responsable d'une ischémie progressive.
Deux zones sont alors identifiables (figure 27.4) :
- la pénombre ischémique : l'oxygénation cérébrale est encore suffisante pour maintenir la viabilité cellulaire, mais l'activité électrique neuronale en revanche ne peut être poursuivie (« silence » électrophysiologique, responsable d'une perte de fonction clinique) ;
- l'infarctus (ou cœur ischémique) : DSC, volume sanguin cérébral et oxygénation sont effondrés, entraînant une mort neuronale avec nécrose tissulaire qui est irréversible.
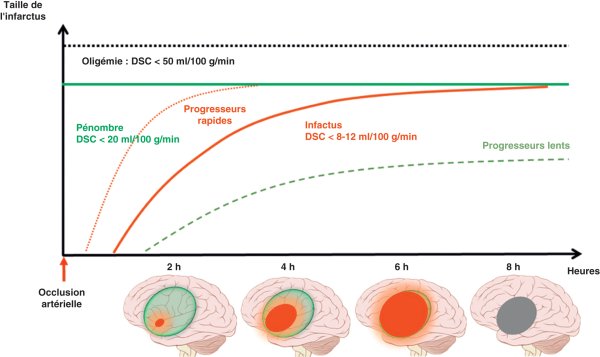
Figure 27.4
Physiopathologie de l'ischémie cérébrale.
Après l'occlusion artérielle, la pénombre s'installe lorsque le débit sanguin cérébral (DSC) est < 20 mL/100 g/min environ. L'évolution vers l'infarctus (nécrose) est plus ou moins rapide selon les individus (progresseurs rapides vs lents), mais intervient lorsque le DSC est < 8-12 mL/100 g/min. La zone infarcie augmente donc progressivement avec le temps d'occlusion.
© Loïc Legris.
La pénombre correspond donc à une zone de souffrance cellulaire pouvant récupérer en cas de reperfusion/réoxygénation suffisamment rapide.
Ainsi, le déficit clinique ne correspond pas uniquement à la zone de nécrose car il inclut le « silence » électrophysiologique des neurones en zone de pénombre. Ceci explique que le déficit constaté soit parfois plus sévère que ce que ne le voudrait l'imagerie, car cette dernière ne permet la visualisation que de la zone nécrosée (en séquence IRM de diffusion notamment).
Cette discordance, aussi appelée mismatch radioclinique, est fondamentale en pratique courante de neurologie vasculaire.
L'infarctus croît aux dépens de la zone de pénombre, et cette évolution est très variable selon la vasoréactivité, les suppléances artérielles, la neuroprotection endogène et un éventuel traitement de reperfusion (recanalisation de l'artère occluse).
Mort cellulaire
Le tissu cérébral est très sensible à la diminution d'apport en O2 et en glucose en raison d'une consommation énergétique élevée avec un métabolisme du glucose quasi exclusivement aérobie et l'absence de réserve in situ. Les mécanismes conduisant à la mort cellulaire impliquent l'ensemble de l'unité neurovasculaire (neurone, astrocyte, cellule endothéliale, péricyte, membrane basale et matrice extracellulaire), les cellules musculaires lisses artériolaires, les oligodendrocytes et la microglie.
L'augmentation du calcium intracellulaire est l'élément déclencheur d'une cascade d'effets délétères qui, mis bout à bout, entraînent la mort neuronale.
L'apoptose correspond à un processus actif, programmé, de la mort cellulaire. L'une des caractéristiques de l'apoptose est liée à la fragmentation de l'ADN (acide désoxyribonucléique) par le biais d'enzymes activées dans le programme apoptotique. Il s'agit d'un mécanisme naturel qui intervient dans l'embryogenèse, mais que l'on retrouve aussi à la suite de lésions du SNC. Elle survient notamment dans la zone de pénombre lorsque la lésion cérébrale est d'origine ischémique et pourrait perdurer plusieurs jours voire semaines après l'AVC. Cibler ce mécanisme pourrait être une piste thérapeutique intéressante dans le futur.
- La vascularisation cérébrale est assurée par le système carotidien et le système vertébrobasilaire.
- Le cercle artériel de la base du cerveau permet l'anastomose entre ces deux systèmes d'une part, et entre le réseau carotidien gauche et droit d'autre part.
- Les variantes du cercle artériel de la base du cerveau sont fréquentes et portent majoritairement sur sa partie postérieure.
- L'autorégulation cérébrale permet de maintenir le débit sanguin cérébral stable face aux variations de la pression artérielle.
- La vasoréactivité cérébrale au CO2, également connue sous le nom de réserve vasomotrice, correspond à la capacité du réseau microcirculatoire artériel de se dilater en réponse au CO2.
- L'AVC ischémique correspond à un arrêt brutal et focal de la perfusion cérébrale. Il s'agit d'une urgence absolue diagnostique et thérapeutique.
- La quantification et la localisation d'une zone dite de « pénombre ischémique » permettent, dans le cas des AVC vus relativement tard, d'évaluer l'efficacité potentielle des traitements de reperfusion cérébrale.
Voir QRM chapitre 32.
Pour en savoir plus
- Baron JC, Marchal G. Ischemic core and penumbra in human stroke. Stroke 1999 ;30:1150–3.
- Deb P, Sharma S, Hassan KM. Pathophysiologic mechanisms of acute ischemic stroke : an overview with emphasis on therapeutic significance beyond thrombolysis. Pathophysiology 2010 ;17:197–218.
- Detante O, Jaillard A, Moisan A, Barbieux M, Favre I, Garambois K, et al. Physiopathologie de l'ischémie cérébrale. EMC Neurologie 2014. 17-045-A-80.
- Dupui P, Géraud G. Régulation de la circulation cérébrale. EMC Neurologie 2006. 17-003-C-10.
- Heiss WD. The ischemic penumbra : how does tissue injury evolve ? Ann N Y Acad Sci 2012 ;1268:26–34.
- Kuschinsky W, Wahl M. Local chemical and neurogenic regulation of cerebral vascular resistance. Physiol Rev 1978 ;58:656–89.
- Miller BA, Bresnahan JC, Beattie MS. Apoptosis in nervous system injury. In: Squire LR, editor. Encyclopedia of neuroscience. Philadelphia: Elsevier ; 2009. p. 523–9.
- Paulson OB, Strandgaard S, Edvinsson L. Cerebral autoregulation. Cerebrovasc Brain Metab Rev 1990 ;2:161–92.
- Tuo QZ, Zhang ST, Lei P. Mechanisms of neuronal cell death in ischemic stroke and their therapeutic implications. Med Res Rev 2022 ;42:259–305.
- Zhang JH, Badaut J, Tang J, Obenaus A, Hartman R, Pearce WJ. The vascular neural network- -a new paradigm in stroke pathophysiology. Nat Rev Neurol 2012 ;8:711–6.