Martin Catala
L'étude du développement embryonnaire et fœtal a connu une véritable révolution depuis quelques années. Celle-ci est largement due au développement de multiples techniques qui permettent de poser des questions fondamentales et d'essayer de soulever le voile des mécanismes qui sous-tendent ces processus. Il est bien entendu exclu dans ce chapitre de traiter l'ensemble du développement du SNC, du SNP et des muscles striés squelettiques.
L'étudiant qui se réfère à ce chapitre est invité à revoir les notions d'embryologie fondamentale telles qu'elles lui ont été présentées au cours de sa 1re année d'études de santé. La nomenclature internationale est utilisée dans ce chapitre. De même, l'étudiant se doit de connaître les notions d'anatomie et d'histologie qui sont un préalable pour appréhender ce chapitre.
À l'issue de la gastrulation, qui se déroule lors de la 3e semaine après la fécondation dans l'espèce humaine, l'embryon se présente sous la forme de trois feuillets primordiaux :
- l'ectoderme est le tissu le plus superficiel. Il se subdivise en deux territoires lors de l'induction neurale, processus complexe non détaillé ici :
- le neurectoderme (précurseur du système nerveux),
- l'ectoderme de surface qui donne naissance à l'épiderme et certaines des annexes cutanées ;
- le mésoderme intermédiaire avec ses domaines (axial, para-axial, intermédiaire et latéral) ;
- l'endoderme profond qui est à l'origine des épithéliums digestifs et respiratoires ainsi que des épithéliums glandulaires associés à ces deux systèmes.
Neurulation
Ce processus morphogénétique rend compte de la transformation de la plaque neurale issue de l'induction neurale en un tube neural. Plusieurs modes de transformation ont été décrits montrant la diversité des phénomènes qui peuvent s'appliquer à ces structures. Il est impossible de tenter d'être exhaustif ici ; néanmoins, il est important de prendre conscience que le modèle proposé est simpliste et la compréhension des dysfonctions de ce temps morphogénétique nécessite une approche pluridisciplinaire. La neurulation primaire est décrite ici dans son aspect le plus simple. Les variantes de la neurulation primaire, les formes secondaire et jonctionnelle ne sont pas abordées car du domaine de l'hyperspécialisation.
La neurulation primaire peut être décrite par la succession de différentes étapes morphogénétiques.
Façonnage
Ce temps morphogénétique rend compte de la déformation de la plaque neurale qui est initialement sous la forme d'un disque et qui se déforme pour prendre un aspect ovoïde (le grand axe est antéropostérieur, c'est-à-dire céphalocaudal ; le petit axe médiolatéral). Ce façonnage est aussi connu sous le terme générique de convergence – extension (convergence vers la ligne médiane et extension antéropostérieure). À l'issue de cette phase (figure 3.1A), le neurectoderme apparaît épaissi et prend le nom de plaque neurale. Le moteur de ce mouvement morphogénétique réside dans les changements de forme des cellules du neurectoderme, dans les mouvements d'intercalation cellulaire ainsi que dans la directionnalité des mitoses. Tout ceci est dicté par la voie Wnt comme pour tous les cas de convergence – extension au cours du développement. Une perturbation génétique de cette voie conduit à un défaut de fermeture du tube neural chez la souris. Des variants de ces gènes pourraient être des facteurs de risque chez les humains.
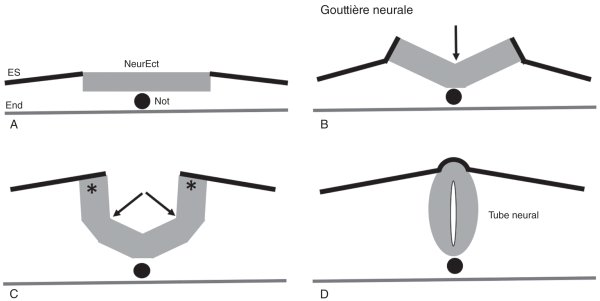
Figure 3.1
Neurulation primaire.
A. Après le façonnage, le neurectoderme (NeurEct) forme un épithélium prismatique et prend le nom de plaque neurale alors que l'ectoderme de surface (ES) est pavimenteux. La notochorde (Not) est située ventralement par rapport à la ligne médiane du neurectoderme. L'endoderme (End) est le tissu le plus ventral de l'embryon à ce stade.
B. Formation de la charnière médiane (flèche). Cette région conduit à la surélévation des bords latéraux de la plaque qui prend alors le nom de gouttière neurale.
C. Formation des deux charnières dorsolatérales (flèches). Les bords de la plaque (bourrelets neuraux) (*) se rapprochent de la ligne médiane où ils fusionneront.
D. À l'issue de la neurulation primaire, le tube neural est formé, il est recouvert par l'ectoderme de surface. Noter qu'aucun espace ne sépare ces deux tissus au niveau de la ligne médiane dorsale.
Plicature
Le premier signe de cette phase morphogénétique est l'apparition d'un sillon médian au niveau de la plaque neurale. Puis, cette plaque se replie lors de la formation de charnières, la première qui se forme est la charnière médiane située dorsalement par rapport à la notochorde (figure 3.1B). La plaque repliée prend alors le nom de gouttière neurale. Puis, deux autres charnières se mettent en place dans les régions dorsolatérales (figure 3.1C). Ceci conduit les régions les plus latérales de la plaque neurale à se rapprocher de la ligne médiane dorsale. Ces régions latérales portent alors le nom de bourrelets neuraux. Le rapprochement définitif de ces bourrelets est assuré par une prolifération cellulaire responsable d'un mouvement de flambage.
Fusion
La plicature conduit au rapprochement des bords latéraux de l'ensemble plaque neurale – ectoderme de surface. À ce stade de développement, ces deux tissus sont fortement adhérents entre eux. Avec le rapprochement sur la ligne médiane dorsale, les deux régions latérales entrent en contact et les tissus homologues fusionnent : le neurectoderme (respectivement l'ectoderme de surface) avec son contingent controlatéral. Ainsi se forme le tube neural recouvert par l'ectoderme de surface (figure 3.1D). À ce stade, il n'existe pas d'espace entre ces deux tissus (figure 3.1D), un tel espace se forme plus tard.
Un tel processus morphogénétique n'est pas uniforme tout au long de l'axe antéropostérieur. Par exemple, l'aspect de la formation du tube neural céphalique est très différent. De plus, chez la souris, il existe des variantes morphologiques selon les grandes régions de la moelle spinale. Rien n'est connu concernant la moelle spinale dans l'espèce humaine du fait de la pauvreté des sources (il est très rare de pouvoir observer des embryons humains de cet âge).
Il est important de constater que les deux lignes médianes du tube neural (en position ventrale et dorsale) n'ont pas du tout la même signification morphogénétique. Ceci suggère que la régulation de leur mise en place est différente et invite à une très grande prudence dans l'utilisation du terme « pathologies de la ligne médiane » pour rendre compte de toutes les malformations siégeant à ce niveau. Ce terme très générique est un « fourre-tout » nosologique qui risque d'entraîner plus de confusion que de solutions pratiques.
Les anomalies de la neurulation primaire entraînent un défaut de fermeture du tube neural qui reste alors exposé à la surface. Si ce défaut touche l'extrémité céphalique, il porte le nom d'anencéphalie ; s'il intéresse la moelle spinale, il se nomme myéloméningocèle ; enfin le craniorachischisis, très rare, touche l'ensemble de l'axe nerveux. Les autres anomalies congénitales sont recouvertes de peau, elles sont liées à des troubles survenant après la neurulation et constituent le groupe des troubles fermés du tube neural.
Développement des muscles striés squelettiques
Seuls les muscles striés squelettiques sont présentés ici. En effet, les muscles lisses et le muscle strié cardiaque représentent des tissus dont la pathologie appartient à d'autres disciplines. Les cellules musculaires striées squelettiques dérivent du mésoderme mis en place lors de la gastrulation. Rappelons que ce feuillet provient de la ligne primitive, les cellules épithéliales de cette ligne perdent leur caractère épithélial pour devenir mésenchymateuses (transition épithéliomésenchymateuse) et migrent entre ectoderme et endoderme. Elles forment alors le troisième feuillet ou mésoderme. Ce mouvement d'internalisation cellulaire prend le nom d'ingression par opposition au mouvement d'invagination (où les cellules s'internalisent en gardant leur caractère épithélial). Selon sa position dans l'axe médiolatéral, le mésoderme se dispose en plusieurs domaines. Les muscles striés squelettiques dérivent essentiellement du domaine para-axial (situé de part et d'autre de la notochorde). L'origine des cellules musculaires striées squelettiques de la tête et de la région ventrale du cou diffère de celle des autres cellules. Le contingent corporel est présenté, puis la région céphalique.
Somites, myotome et muscles striés squelettiques du corps
Le mésoderme para-axial issu de la ligne primitive se met en place selon un gradient rostrocaudal (les cellules les plus caudales s'ajoutent à l'extrémité déjà formée selon le mode de croissance appelé accrétion). Il forme alors le mésoderme présomitique dont l'extrémité rostrale se condense pour former un cube ou somite (figure 3.2). Cette structure mésodermique est constituée par un mur épithélial entourant un cœur mésenchymateux. Le somite subit l'action polarisatrice des tissus environnants et donne naissance au sclérotome ventral et au dermomyotome dorsal, ce dernier se divisant en dermatome situé sous l'ectoderme de surface et en myotome intermédiaire (figure 3.2). Les cellules musculaires striées squelettiques du corps proviennent du myotome à l'exception des muscles sterno-cléido-mastoïdien et trapèze dont les cellules musculaires proviennent des lames latérales cervicales. Les myotomes génèrent des cellules mononucléées (les myoblastes) qui migrent pour atteindre leur lieu définitif de différenciation. Là, ils subissent une maturation caractérisée par leur fusion générant des myotubes (dont les noyaux occupent une position encore centrale). La maturation terminale conduit au déplacement des noyaux en périphérie et à la maturation de la jonction entre le motoneurone périphérique et le muscle (jonction neuromusculaire). Toutes ces phases sont parfaitement régulées par des systèmes moléculaires bien connus, non approfondis dans ce chapitre.
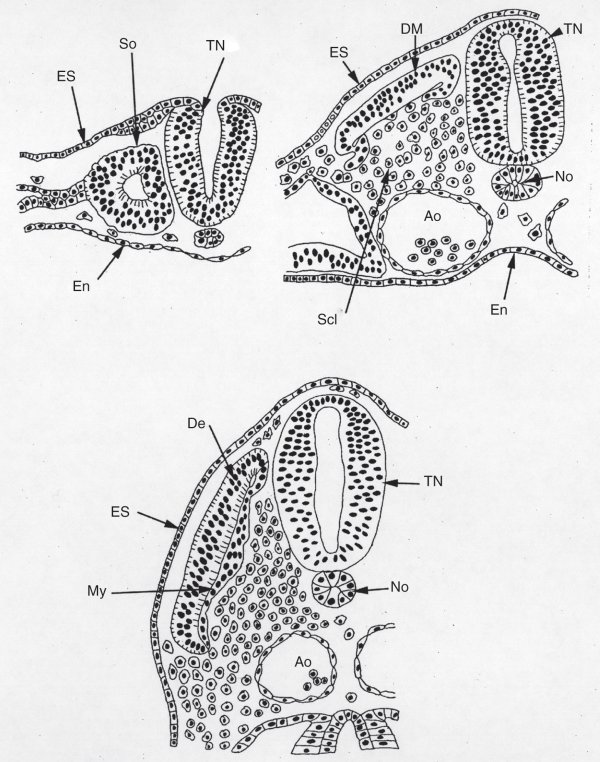
Figure 3.2
Coupes transversales d'un embryon montrant la maturation du somite.
Le somite (So) résulte de la segmentation du mésoderme présomitique mis en place lors de la gastrulation. Il se dissocie en dermomyotome (DM) dorsal et sclérotome (Scl) ventral. Puis le dermomyotome évolue pour générer le dermatome (De) et le myotome (My) qui donne naissance aux cellules musculaires striées squelettiques. Ao : aorte, En : endoderme, ES : ectoderme de surface, No : notochorde, TN : tube neural.
© Catala M. Embryologie. Développement précoce chez l'humain, 3e éd. Paris : Elsevier Masson ; 2011.
Il convient de distinguer les muscles axiaux (ou domaine épaxial), dont les cellules musculaires proviennent du myotome médian (au contact du tube neural), et les muscles latéroventraux (y compris les muscles des membres) (ou domaine hypaxial), dont les cellules musculaires proviennent des myotomes latéraux.
Par ailleurs, les tendons et les cellules conjonctives du muscle strié squelettique n'ont pas la même origine que les cellules musculaires. Les cellules de ces composants proviennent d'une région différente du somite s'il s'agit du domaine épaxial et du mésoderme somatopleural s'il s'agit du domaine hypaxial.
Enfin, la forme du muscle dépend non pas de l'origine des cellules musculaires striées squelettiques mais de celle des cellules conjonctives. Ainsi, les variations anatomiques de la forme des muscles dépendent donc des cellules conjonctives et ne sont pas explicables par des anomalies des cellules musculaires elles-mêmes.
Muscles striés squelettiques de la tête et de la région ventrale du cou
Les cellules musculaires striées squelettiques de ces régions ne proviennent pas des somites car cette partie de l'embryon en est dépourvue. Elles proviennent du mésoderme para-axial rostral non segmenté (ou mésoderme céphalique) situé dans le prolongement des somites mais aussi de l'extrémité la plus rostrale du mésoderme axial (ou plaque préchordale). L'origine muscle par muscle n'est pas détaillée dans ce chapitre. Il convient néanmoins de préciser qu'il s'agit des muscles oculomoteurs extrinsèques, des muscles faciaux et des muscles issus des arcs pharyngiens (région ventrale du cou).
Développement du système nerveux périphérique
Le système nerveux périphérique est défini anatomiquement et histologiquement par les structures situées en dehors des centres nerveux. Les cellules gliales (cellules de Schwann et cellules satellites des ganglions nerveux) qui composent le SNP sont radicalement différentes de celles qui constituent le SNC (astrocytes, oligodendrocytes et cellules microgliales) (cf. chapitre 2). Néanmoins, il est important de noter que les axones périphériques peuvent provenir de cellules du SNC (ex : motoneurones) ou que des axones de cellules périphériques peuvent cheminer dans le SNC (ex : axones sensitifs issus des cellules des ganglions de la racine dorsale). Ainsi des anomalies des régions centrales peuvent donner des signes cliniques généralement associés à des anomalies du SNP (l'exemple typique est l'atteinte de la corne ventrale de la moelle spinale qui génère un déficit moteur de type périphérique). Le deuxième point important à connaître réside dans la différence fondamentale entre les SNP spinal et crânien.
Système nerveux périphérique spinal
Il convient de séparer les neurones périphériques de projection (motoneurones périphériques et neurones végétatifs spinaux) et les cellules des ganglions périphériques (ganglions de la racine dorsale, ganglions végétatifs). Les motoneurones périphériques, comme les neurones végétatifs de projection issus de la moelle spinale, naissent dans le tube neural. Leurs axones grandissent et quittent le tube, ils forment alors la racine ventrale des nerfs spinaux. Ces axones ne peuvent migrer qu'au niveau du sclérotome issu de l'hémisomite rostral. Le sclérotome issu de l'hémisomite caudal est imperméable pour la croissance des axones. De ce fait, les neurones de projection du système nerveux périphérique, même s'ils sont produits tout au long de l'axe antéropostérieur, forment des racines distinctes séparées les unes des autres. La segmentation des racines est donc imposée par l'environnement somitique. Au-delà du sclérotome rostral, les axones grandissent en suivant les voies de migration déterminées par la matrice extracellulaire. Ainsi, la forme des plexus nerveux issus des racines dépend de cette matrice générée par le mésoderme somatopleural. De ce fait, la morphologie générale du SNP spinal dépend des tissus non neuraux qui permettent la pousse axonale. Les variations anatomiques des plexus et des nerfs (ex : anastomoses) s'expliquent donc par la propriété de l'environnement et non des axones nerveux.
Les ganglions du SNP sont générés par des cellules issues du toit du tube neural. Ce toit subit une transformation radicale : certaines de ses cellules s'engagent dans un processus de transition épithéliomésenchymateuse (elles perdent leur caractère épithélial pour devenir mésenchymateuses et elles s'individualisent isolément entre l'ectoderme de surface et le tube neural) (figure 3.3). Ces cellules isolées forment la crête neurale qui peut alors migrer suivant trois voies de migration :
- la voie la plus dorsale permet la mise en place des mélanocytes (qui ne sont pas un constituant du SNP) ;
- la voie qui traverse les dérivés de l'hémisomite rostral conduit les cellules de la crête neurale à s'agréger pour former le ganglion de la racine dorsale (ou ganglion spinal) ;
- la voie ventrale assure la migration des cellules végétatives (figure 3.3).
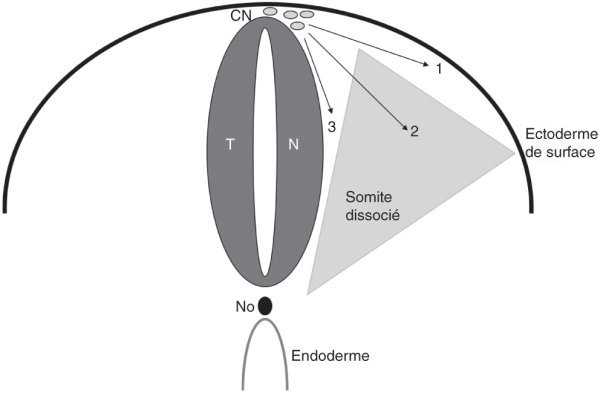
Figure 3.3
Naissance des cellules de la crête neurale (CN) depuis la région la plus dorsale ou toit du tube neural (TN).
Elles peuvent migrer selon trois voies : la voie sous-ectodermique (1) permet la migration des mélanocytes, la voie somitique (2) conduit les cellules à s'agréger pour former le ganglion de la racine dorsale, et la voie ventrale (3) permet la migration des neurones végétatifs. No : notochorde.
De la même manière que pour les racines ventrales, la perméabilité différentielle entre sclérotome rostral et caudal conduit à la segmentation des racines dorsales et des ganglions spinaux.
Système nerveux périphérique crânien
La situation est de loin plus complexe en ce qui concerne les nerfs crâniens. La crête neurale des régions crâniennes est une structure qui participe non seulement à la genèse du SNP mais aussi à la genèse d'une partie du squelette de la face et de la région ventrale du cou. De plus, le SNP de ces régions ne dérive pas exclusivement de la crête neurale contrairement au SNP spinal. En effet, certains neurones ou cellules de soutien de ganglions des nerfs crâniens sont produits à partir d'épaississements de l'ectoderme de surface, dénommés placodes. Par exemple, la placode otique génère, entre autres dérivés, les neurones des ganglions auditifs et vestibulaires de la VIIIe paire de nerfs crâniens. L'origine précise des différentes structures du SNP de cette région n'est pas développée davantage dans ce chapitre.
Développement du cervelet
Le cervelet dérive du tube neural et plus précisément du premier rhombomère (c.-à-d. la région la plus rostrale du métencéphale). Il est aujourd'hui clairement établi, tant chez les oiseaux que chez les rongeurs, que le cervelet est initialement présent sous la forme de deux ébauches séparées par la ligne médiodorsale (figure 3.4A). Il est à noter que la polarité antéropostérieure initiale de ces ébauches correspond à la future polarité médiolatérale du cervelet. Le tube neural de la région rhombomérique se déforme à la suite du développement de l'ébauche du 4e ventricule qui prend la forme d'un losange. Cette déformation conduit à une bascule des ébauches cérébelleuses (figure 3.4B), leurs régions initialement rostrales devenant plus médianes. L'évolution de ce mouvement morphogénétique conduit à la fusion des régions médianes (figure 3.4C). À l'issue de ce temps morphogénétique, le rhombencéphale apparaît losangique en vue dorsale. Ce losange représente le futur 4e ventricule, son toit est constitué d'un tissu épithélial très fin. Les bords du losange sont épais, blanchâtres et constituent les lèvres rhombiques (avec un contingent rostral dont le cervelet est issu et un contingent caudal).
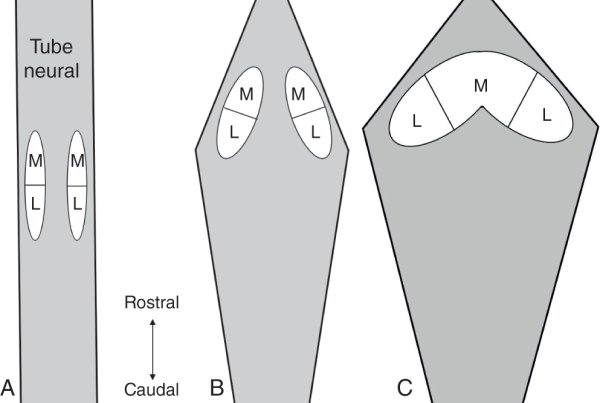
Figure 3.4
Origine précoce du cervelet dans le tube neural.
A. Initialement, deux ébauches cérébelleuses sont présentes dans le tube neural. La région rostrale de ces ébauches donne naissance à la future région médiane du cervelet (M) alors que la région caudale participe à la formation des régions latérales (L). B. Avec la déformation de la région rhombencéphalique secondaire à la croissance de l'ébauche du 4e ventricule, l'axe rostrocaudal initial change et prend une direction médiolatérale. C. La croissance du ventricule conduit ensuite à la fusion des ébauches médianes (le tissu médian est chassé vers les régions rostrale et caudale du tube). Ainsi se constitue l'axe définitif de l'ébauche cérébelleuse.
Après fusion des deux ébauches, on distingue deux zones générant des neurones dans le primordium cérébelleux : la zone ventriculaire et la lèvre rhombique (figure 3.5A). La zone ventriculaire donne naissance aux cellules inhibitrices du cervelet (dans les noyaux profonds ainsi que dans le cortex cérébelleux) (figure 3.5B). Dans le cortex cérébelleux, de nombreux interneurones sont inhibiteurs. Il en est de même des cellules de Purkinje. La lèvre rhombique produit quant à elle tous les neurones excitateurs (des noyaux profonds comme du cortex cérébelleux). Dans un premier temps, les cellules migrent à la surface de l'ébauche cérébelleuse formant une couche transitoire (la couche granulaire externe). Puis les cellules de cette couche se différencient et migrent en rayons vers le ventricule pour former la couche des grains internes (ou couche granulaire interne du cervelet). Dans l'espèce humaine, cette maturation s'étend après la naissance si bien qu'une neurogenèse se poursuit tardivement dans cette région.
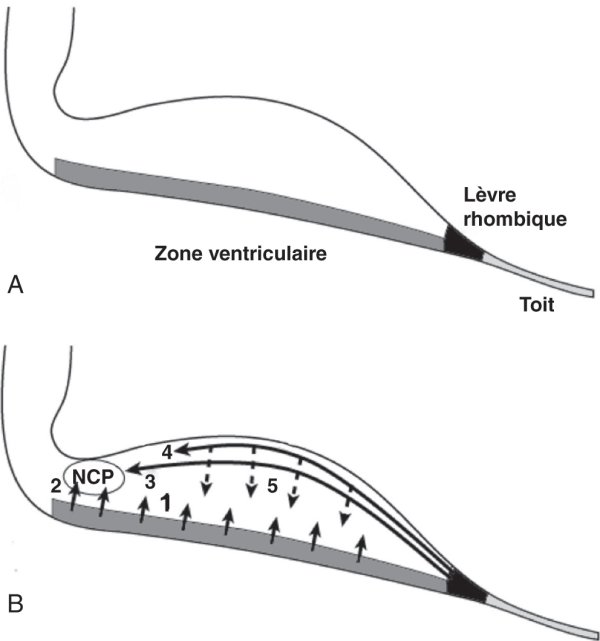
Figure 3.5
Mouvements cellulaires lors de la morphogenèse du cervelet.
A. Coupe sagittale de l'ébauche cérébelleuse. La zone ventriculaire est située au contact du 4e ventricule. La lèvre rhombique forme l'interface entre l'ébauche cérébelleuse et le toit du 4e ventricule. B. À partir de la zone ventriculaire, des cellules migrent de manière radiaire pour former les cellules inhibitrices du cortex cérébelleux dont les cellules de Purkinje (1) et les cellules inhibitrices des noyaux profonds du cervelet (NCP) (2). Les cellules issues de la lèvre rhombique migrent superficiellement pour générer les cellules excitatrices des NCP (3) ainsi que les cellules qui forment la couche granulaire externe (4). À partir de cette dernière couche, les cellules migrent en rayons pour générer les cellules excitatrices du cortex cérébelleux dont les cellules de la couche des grains (5).
Des fissures se creusent progressivement dans l'ébauche cérébelleuse de façon stéréotypée. Ces structures sont à la base de la segmentation cérébelleuse caractérisée par une foliation. La génération de telles fissures est encore très mal comprise : il existe des zones de prolifération cellulaire associée à des modifications des contraintes mécaniques qui conduisent à la formation de points d'ancrage, précurseurs de la région profonde des futurs folia.
De nombreuses malformations peuvent affecter le cervelet. Le terme d'atrophie cérébelleuse suppose que le cervelet s'est formé normalement et que son volume régresse secondairement. Ce n'est donc pas une malformation à proprement parler. Si le cervelet est de petite taille dès sa formation, on parle alors d'hypoplasie. Enfin, si les folia sont désorganisés entraînant une anomalie de la forme du cervelet, on parle de dysplasie.
Développement de l'isocortex cérébral
Avant de commencer la description de la formation du cortex cérébral, il semble important de noter quelques discordances entre anatomistes et biologistes du développement. Les hémisphères cérébraux sont des organes creux centrés sur les ventricules cérébraux et, par convention pour les anatomistes, la région des ventricules est considérée comme la partie interne ou profonde, la région sous-piale comme la zone la plus externe ou superficielle. Au contraire, pour les spécialistes du développement, l'origine de ces régions est le tube neural qui se présente comme un épithélium. Or, la région basale de cet épithélium est située au niveau anatomique superficiel alors que la région apicale du tube est ventriculaire. Il est donc important de noter cette discordance qui pourrait entraîner des confusions.
Les mécanismes qui conduisent à la formation du cortex cérébral ont été entièrement repensés ces 30 dernières années du fait des connaissances nouvelles acquises en particulier chez les rongeurs. Tous les modèles développés avant 1990 sont désormais obsolètes et n'ont plus qu'un intérêt historique. Le cortex cérébral humain est essentiellement un isocortex et ne sont présentées ici que les données concernant ce type de cortex. La région corticale provient des vésicules télencéphaliques qui sont des dérivés dorsaux du tube neural. Ces vésicules sont subdivisées en deux grandes régions : le pallium (la région la plus dorsale) et le sous-pallium (région intermédiaire qui donne naissance aux éminences ganglionnaires).
Le pallium initialement se présente sous la forme d'un neuroépithélium comme l'ensemble du tube neural. À partir des régions médiane (l'ourlet cortical) et latérale (la frontière pallium – sous-pallium) (figure 3.6A), des cellules migrent en superficie (c.-à-d. entre le neuroépithélium et le tissu conjonctif qui l'enveloppe) pour recouvrir l'ensemble de la surface télencéphalique. Cette couche forme la préplaque (figure 3.7A), couche pauvre en cellules et riche en prolongements qui s'enchevêtrent sous la forme d'un plexus.
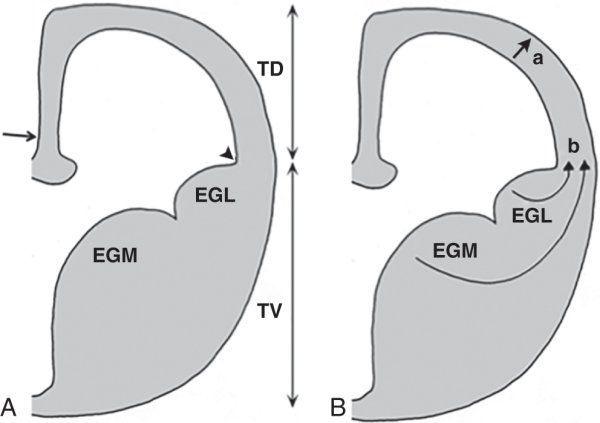
Figure 3.6
Coupes coronales du télencéphale au cours du développement.
A. On subdivise le télencéphale en une région dorsale (TD) et une région ventrale (TV). La région ventrale contient des éminences ganglionnaires (médiane : EGM, et latérale : EGL). La limite entre les deux zones télencéphaliques (tête de flèche) est appelée frontière pallium – sous-pallium. L'ourlet cortical (flèche) se situe au niveau médian. La préplaque naît de ces deux régions. B. Les neurones formant la plaque corticale ont une double origine : les neurones de projection (pyramidaux) proviennent de l'épithélium ventriculaire ou sous-ventriculaire (a) alors que les interneurones corticaux naissent dans les éminences ganglionnaires (b).
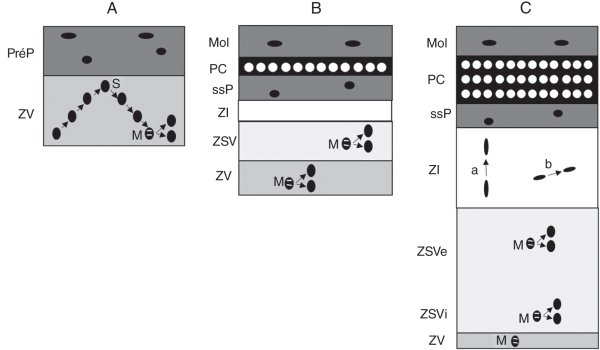
Figure 3.7
Phases du développement de l'isocortex cérébral.
A. Les cellules de la préplaque (PréP) proviennent de l'ourlet cortical et de la frontière pallium – sous-pallium. Elles recouvrent l'épithélium ventriculaire ou zone ventriculaire (ZV) où se situent les mitoses (M). B. Les neurones qui se différencient migrent et scindent la PréP en deux sous-régions : la couche moléculaire (Mol) et la sous-plaque (ssP). Ces neurones forment une nouvelle couche, la plaque corticale (PC). La zone ventriculaire (ZV) est recouverte d'une couche sous-ventriculaire (ZSV). Les mitoses sont observées dans la ZV et la ZSV. La phase de synthèse de la mitose (S) se déroule à la jonction ZV-PréP. La zone intermédiaire (ZI) est le lieu de migration des cellules nouvellement différenciées. C. La zone sous-ventriculaire s'étend avec deux composantes interne (ZSVi) et externe (ZSVe). Dans la ZI, on observe des migrations radiaire (a) ou tangentielle (b). La plaque corticale s'épaissit au fur et à mesure du développement (gradient interne – externe).
Après la formation de la préplaque, les cellules neuronales qui se différencient migrent à travers la zone intermédiaire et divisent cette préplaque en deux régions : la zone moléculaire (en superficie, soit sous la méninge primitive) et la sous-plaque (figure 3.7B). Au niveau ventriculaire, une deuxième couche se différencie (la zone sous-ventriculaire). Les cellules qui génèrent la plaque corticale ont une double origine (figure 3.6B) : les cellules pyramidales (de projection) proviennent des régions ventriculaire et sous-ventriculaire du télencéphale dorsal alors que les interneurones corticaux sont issus des éminences ganglionnaires. Au cours de l'évolution de ces structures, la couche sous-ventriculaire s'étend considérablement, formant deux sous-régions : la couche sous-ventriculaire interne et la couche sous-ventriculaire externe. Les mitoses générant les neurones de projection surviennent dans la couche ventriculaire et les deux composantes de la couche sous-ventriculaire. Les cellules migrent selon différents modes : radiaire et tangentiel. Une même cellule peut suivre un trajet initialement radiaire puis changer de direction. Finalement, la cellule néoformée traverse la sous-plaque et toutes les couches déjà établies de la plaque corticale. Elle se dispose alors entre la plaque corticale et la couche moléculaire. Ce mode de génération est appelé gradient interne – externe (pour rendre compte de la différenciation plus tardive des cellules plus superficielles). La couche I de l'isocortex provient de la couche moléculaire, la plaque corticale génère les couches II à VI et la sous-plaque disparaît quasi totalement.
Enfin, les sillons corticaux se développent plus tardivement. Ils se mettent en place progressivement, débutant par des sillons constants et se poursuivant par des sillons beaucoup plus variables. Le mécanisme exact de leur mise en place est encore très largement discuté.
De nombreuses anomalies du développement peuvent être observées chez l'être humain. De façon schématique, on peut décrire des neurones situés dans une position aberrante ou hétérotopies qui peuvent siéger en sous-ventriculaire, dans la substance blanche ou dans les espaces méningés. Les anomalies de forme ou d'organisation des sillons sont représentées par les lissencéphalies I et II, les polymicrogyries, etc. Il existe de très nombreuses anomalies corticales qui sont du domaine de l'hyperspécialisation.
Voir QRM chapitre 32.