Xavier MoissLénaic Monconduit et Nadine Attal
La douleur est définie par l'International Association for the Study of Pain (IASP) comme « une expérience sensorielle et émotionnelle désagréable, associée ou semblant être associée à une lésion tissulaire réelle ou potentielle » (définition mise à jour en 2020). Cette définition indique qu'une douleur est à la fois une sensation et une émotion, ce qui implique différents modes de transmission anatomophysiologiques.
La douleur est un symptôme clinique très fréquent et un motif de consultation médicale quotidien. Si la douleur aiguë constitue un système d'alerte indispensable, permettant de limiter le risque d'atteinte à l'intégrité de l'organisme, la douleur chronique est considérée comme une maladie à part entière, puisqu'elle perd cette fonction d'alerte et devient source de stress chronique, altérant la qualité de vie du patient.
L'objectif de ce bref chapitre est de décrire la physiologie de la transmission du message douloureux, depuis l'activation des récepteurs périphériques jusqu'à l'intégration de la douleur au niveau cortical, les médiateurs chimiques impliqués dans la transmission de l'influx douloureux, et les principaux systèmes de modulation de la douleur afin de mieux comprendre leurs conséquences en termes d'expérience douloureuse.
Neurones périphériques
S'il existe des récepteurs spécifiques des différentes modalités de la sensibilité épicritique qui regroupe la perception du tact fin et conscient et la proprioception consciente (récepteurs de la vibration, pression, étirement), ce n'est pas le cas pour la sensibilité protopathique qui regroupe la perception de la douleur, de la température et du tact grossier. En effet, les récepteurs de la douleur correspondent aux terminaisons des fibres nerveuses sensitives (aussi appelées neurones de 1er ordre ou protoneurones) de petit calibre appelées Aδ et C (figure 30.1). Ces fibres sont de diamètre beaucoup plus petit que celles véhiculant les informations correspondant à la sensibilité épicritique. Les fibres C ne sont pas myélinisées (amyéliniques), d'où une vitesse de transmission de l'information très lente (de l'ordre de 1 à 2 m/s). Les fibres Aδ, faiblement myélinisées, sont caractérisées par une transmission de l'information plus rapide (de l'ordre de 10 à 20 m/s), mais qui reste plus lente que celle des grosses fibres myélinisées conduisant la sensibilité épicritique (fibres Aβ, conduction à 50-60 m/s). Les fibres sensitives, qu'elles véhiculent une information non douloureuse ou douloureuse, ont toutes leur corps cellulaire au sein du ganglion de la racine dorsale pour les nerfs spinaux et au niveau du ganglion trigéminal (anciennement de Gasser) pour le nerf trijumeau, qui traite les sensations de la face. Toutes ces fibres projettent au niveau de la corne dorsale de la moelle spinale (ou tronc cérébral pour le nerf trijumeau). À ce niveau, il existe une organisation anatomique et électrophysiologique permettant de définir des couches bien distinctes. Les fibres Aδ et C se terminent dans les couches superficielles de la moelle spinale (anciennement moelle épinière), tandis que les fibres Aβ se terminent dans les couches profondes.
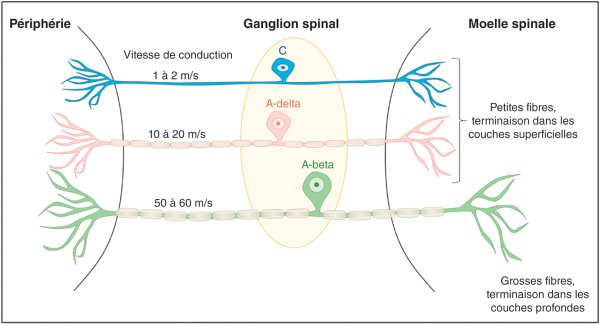
Figure 30.1
Représentation schématique des neurones de 1er ordre.
Les terminaisons nerveuses libres des fibres Aδ et C se situent au niveau de l'épiderme, mais également dans les muqueuses et organes internes (viscères, muscles, articulations, méninges, etc.). Elles peuvent être activées par différents types de stimulations mettant en jeu le système nociceptif :
- le chaud : le seuil d'activation des récepteurs est de 43 °C pour la majorité des fibres. L'un des récepteurs principaux pour la réponse au chaud est le récepteur TRPV1 (transient receptor potential vanilloid type 1), qui est activé par la capsaïcine (composant actif du piment) et constitue une cible thérapeutique pour certaines douleurs neuropathiques ;
- le froid : le seuil d'activation des récepteurs est de 25 °C pour la majorité des fibres. L'un des récepteurs principaux pour la réponse au froid est le récepteur TRPM8 (transient receptor potential menthol type 8), qui est activé notamment par le menthol ;
- les stimulations mécaniques : de nombreux récepteurs semblent impliqués dans la mécanotransduction, notamment des canaux ASIC (acid-sensing ion channels) et TRP (transient receptor potential). En cas de stimulation mécanique intense, il existe une sensation de double douleur :
- la première est ressentie plutôt comme une piqûre localisée, apparaissant de façon quasi instantanée et correspondant à l'activation des fibres Aδ,
- la seconde, plus tardive, perçue avec un décalage qui peut atteindre une seconde, est ressentie comme une brûlure plus diffuse, correspondant à l'activation des fibres C.
Ce décalage temporel dans la perception des deux types de sensation peut notamment être expérimenté lors d'un coup de marteau reçu sur un doigt ou lorsqu'un orteil heurte le pied du lit !
- les stimulations chimiques : là encore, la famille des TRP joue un rôle important, notamment le récepteur TRPA1 (transient receptor potential ankyrin type 1), activé entre autres par des molécules irritantes contenues dans l'ail, le wasabi ou le gaz lacrymogène.
Dès le premier neurone, l'intensité de la douleur est corrélée aux potentiels d'action. En effet, plus la stimulation est intense au niveau des terminaisons nerveuses libres, plus la dépolarisation est importante au niveau neuronal (potentiel générateur), et plus les signaux électriques envoyés via l'axone, sous forme de potentiels d'action, sont rapprochés (figure 30.2). En effet, chaque potentiel d'action ayant une amplitude fixe, l'intensité n'est pas codée en amplitude, mais uniquement en fréquence. Par ailleurs, pour une même intensité de stimulation, le ressenti est différent selon la surface concernée par la stimulation nociceptive : plus celle-ci est large, plus le nombre de récepteurs activés est important. Ainsi, il est tout à fait supportable de plonger un doigt dans de l'eau à 2 °C, mais franchement douloureux d'y plonger un bras entier. Ce phénomène correspond à de la sommation spatiale.
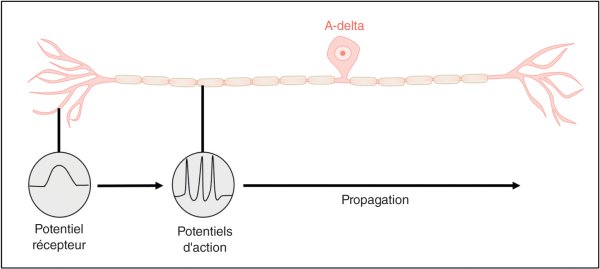
Figure 30.2
Représentation schématique d'un neurone A-delta avec potentiel récepteur
dépendant de la stimulation périphérique et potentiels d'action transmis le long de l'axone.
iUn seul neurone (pouvant aller jusqu'à 1 m de long) couvre l'ensemble de la distance entre l'extrémité de la cellule (terminaisons nerveuses libres situées dans la peau ou les organes internes) et la moelle spinale. Les fibres de petit calibre font synapse au niveau de la corne dorsale de la moelle spinale pour donner naissance au faisceau spinothalamique. Des neurones à convergence sont retrouvés dans les couches plus profondes de la corne dorsale (couche V). Ces neurones reçoivent des afférences nociceptives et non nociceptives provenant de zones cutanées et viscérales, ce qui permet de comprendre l'existence de douleurs projetées.
Exemple
Lésion hépatique dont la douleur est parfois ressentie au niveau de l'épaule droite.
Les lésions tissulaires entraînent la libération de nombreuses substances qui activent ou sensibilisent les nocicepteurs : potassium, ions H+ (acidité), bradykinine, histamine, sérotonine, prostaglandines, leucotriènes, etc. Les fibres nociceptives peuvent libérer en périphérie des neurotransmetteurs classiques (en particulier du glutamate) ainsi que de petits peptides (substance P, CGRP [calcitonin gene-related peptide]) qui ont une action vasodilatatrice, et favoriser la libération d'histamine ou de sérotonine, « sensibilisant » ainsi les nocicepteurs voisins. Ce phénomène correspond à l'inflammation neurogène, à l'origine de l'hyperalgésie primaire (hyperalgésie : douleur en réponse à une stimulation normalement peu douloureuse). Certains antalgiques comme les AINS agissent sur la synthèse des prostaglandines en inhibant l'action de la cyclo-oxygénase (COX). Les corticoïdes agissent sur la même voie mais plus en amont. La lésion tissulaire entraîne également la sécrétion de substances antalgiques comme des peptides opioïdes qui ont ici une action périphérique.
Voies spinothalamiques et principaux neuromédiateurs de la douleur
La voie néospinothalamique, habituellement appelée simplement « spinothalamique » (qui est l'une des voies extralemniscales), véhicule les informations douloureuses et thermiques, par opposition à la voie lemniscale (appelée ainsi car le 2e neurone croise la ligne médiane dans le bulbe, au sein d'une structure appelée lemnisque médian), dans laquelle sont véhiculées les informations du tact non douloureux (figures 30.3 et 30.4). Seules les voies spinothalamiques sont détaillées ici, la sensibilité non douloureuse étant abordée chapitre 7.
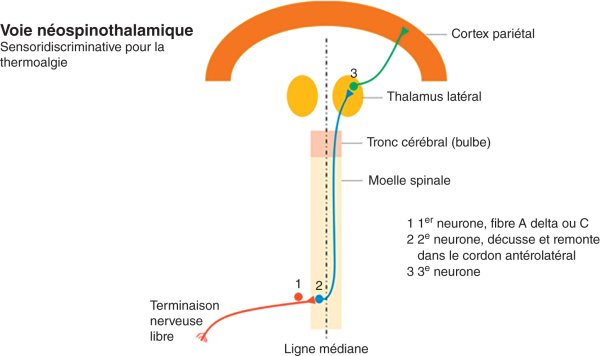
Figure 30.3
Voie néospinothalamique (l'une des voies extralemniscales).
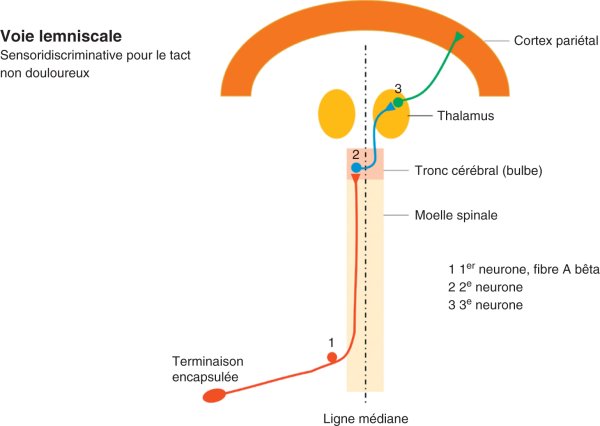
Figure 30.4
Voie lemniscale.
Ces neurones de 2e ordre décussent (franchissent la ligne médiane) et remontent dans le cordon latéral (ou antérolatéral) de la moelle spinale. Ils remontent jusqu'au niveau du thalamus latéral. Une deuxième synapse a lieu au niveau thalamique avec le 3e neurone qui va du thalamus au cortex pariétal. Au niveau du cortex pariétal, il existe une somatotopie (représentation des différentes parties du corps au niveau du système nerveux) très précise. La représentation de chaque zone du corps occupe une place spécifique et reproductible d'un individu à l'autre, au niveau du gyrus postcentral (en arrière du sillon central). La première description très précise de cette somatotopie chez l'homme a été faite par Wilder Penfield, c'est pourquoi on la dénomme souvent homunculus de Penfield.
Les principaux neurotransmetteurs intervenant entre les afférences nociceptives (1ers neurones de la nociception) et les neurones spinaux (2es neurones) sont des neurotransmetteurs classiques (acides aminés excitateurs comme le glutamate ou l'aspartate) et des neuropeptides (substance P, VIP [vasoactive intestinal peptide], CGRP, etc.). Ces neuromédiateurs sont responsables de la transmission du message douloureux mais aussi de phénomènes de « sensibilisation centrale » correspondant à une hyperexcitabilité neuronale dans la moelle spinale ou le cerveau, pouvant rendre compte de l'hyperalgésie secondaire. Cette hyperalgésie secondaire explique notamment l'extension possible d'une zone douloureuse au-delà de la zone initialement lésée.
En résumé
La voie néospinothalamique correspond à la voie sensoridiscriminative qui véhicule les informations nociceptives. De façon physiologique, elle permet d'identifier la localisation de la stimulation qui peut menacer l'intégrité de l'organisme, d'en connaître la nature (piqûre, brûlure, etc.) et l'intensité. Cette voie n'est pas responsable de la part émotionnelle et du ressenti désagréable correspondant à l'expérience douloureuse dans son ensemble.
Autres voies de transmission impliquées dans l'expérience douloureuse
Outre la voie néospinothalamique, de nombreuses autres voies de transmission sont impliquées dans l'expérience douloureuse. Les 2es neurones de la voie paléospinothalamique projettent vers le thalamus médian. Ce dernier, qui reçoit également des informations issues du tronc cérébral, a des projections plus étendues au niveau cortical et notamment sur des aires impliquées dans l'intégration de l'aspect émotionnel (caractère désagréable) de la douleur comme le cortex préfrontal ou le cortex cingulaire. Ces régions corticales jouent notamment un rôle dans la réaction émotionnelle et comportementale à la douleur, visant à permettre à l'individu de réagir face à la douleur (fuir, combattre, etc.). L'amygdale est aussi particulièrement impliquée dans la réaction affective et de colère à la douleur via la voie spino-parabrachio-amygdalienne (le noyau parabrachial étant situé dans la partie dorsale du pont). Une « mise en mémoire » de l'information douloureuse et aversive, dont la structure clé est l'hippocampe, qui joue un rôle crucial dans la mémoire, est également capitale pour éviter de reproduire une situation qui serait vécue comme dangereuse pour l'intégrité de l'organisme. D'autres systèmes, tels que les systèmes spino-hypothalamique et spino-parabrachio-hypothalamique sont à l'origine de la réaction végétative (du système nerveux autonome) à la douleur, qui se traduit par une augmentation de la fréquence cardiaque et de la pression artérielle, pouvant être observée lors d'une douleur aiguë. Ces paramètres cardiovasculaires (pression artérielle, fréquence cardiaque) sont classiquement monitorés par les anesthésistes pour adapter l'antalgie lors d'une chirurgie.
Systèmes de contrôle de la nociception
Contrôles inhibiteurs segmentaires au niveau de la corne dorsale de la moelle spinale
Il a été montré de longue date que les fibres de gros calibre (Aβ, véhiculant la sensibilité non douloureuse) exercent une inhibition sur le faisceau spinothalamique par l'intermédiaire d'interneurones, fermant ainsi la « porte » à la transmission de la douleur (figure 30.5). Ce système de contrôle est lui-même soumis à l'influence de contrôles descendants qui peuvent avoir un effet facilitateur ou inhibiteur sur la transmission du message douloureux. Cette théorie appelée théorie du gate control, ou théorie du portillon est notamment utile pour la compréhension de l'effet antalgique de la neurostimulation transcutanée (TENS) ou de la stimulation médullaire. Elle rend compte de façon simple du mécanisme par lequel une stimulation non douloureuse peut réduire la transmission d'un message douloureux.
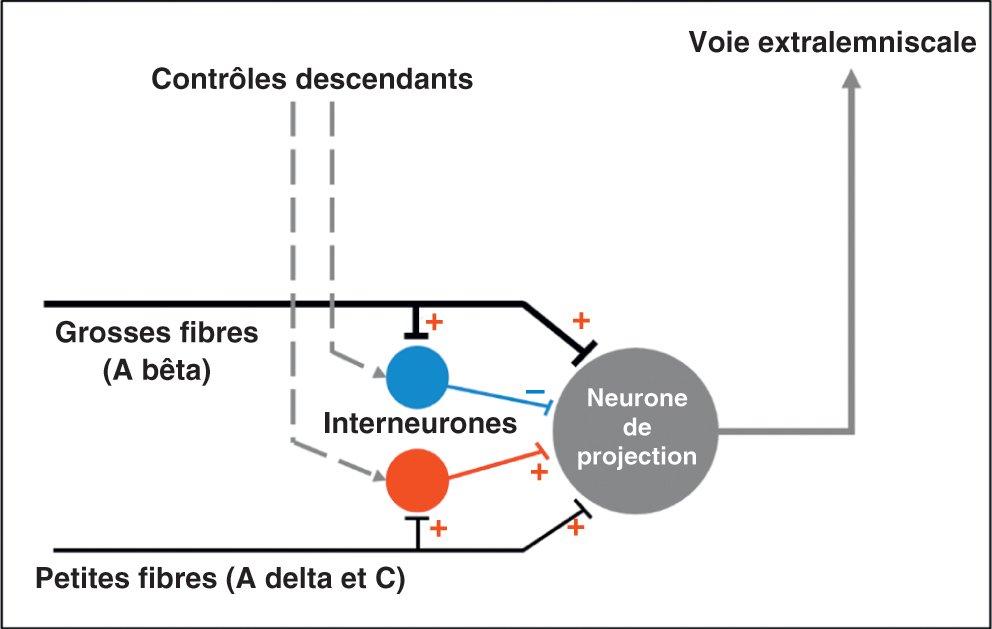
Figure 30.5
Schéma simplifié de la théorie du gate control.
Au niveau médullaire, les interneurones inhibiteurs sont généralement gabaergiques, c'est-à-dire que leur neurotransmetteur est l'acide gamma-aminobutyrique, acide aminé inhibiteur.
Contrôles descendants
Les mécanismes chimiques de la transmission douloureuse au niveau supramédullaire sont plus complexes. Signalons notamment l'existence de nombreux récepteurs opioïdes au niveau du système nerveux central. La sérotonine, la dopamine et la noradrénaline sont largement impliquées dans la régulation de la douleur via des contrôles descendants.
Ces contrôles correspondent à des voies descendantes qui trouvent leur origine au sein de plusieurs structures du tronc cérébral (substance grise périaqueducale, locus coeruleus, raphé magnus, etc.), mais aussi de l'hypothalamus, des noyaux thalamiques ou du cortex (frontal et limbique). Ces voies se projettent dans la moelle spinale avec un rôle facilitateur ou inhibiteur sur les neurones à convergence, en fonction des récepteurs qu'ils activent (noradrénergiques, sérotoninergiques, dopaminergiques, opioïdergiques, etc.).
Contrôles inhibiteurs diffus induits par une stimulation nociceptive
Il existe également des contrôles uniquement inhibiteurs. En effet, le déclenchement d'une douleur focale peut activer des voies de contrôles inhibitrices permettant de réduire l'activité basale des neurones nociceptifs extérieurs à la zone douloureuse. Ce mécanisme, qui rend compte de l'inhibition d'une douleur par une autre douleur, est souvent désigné en clinique sous le terme de « modulation conditionnée de la douleur » (conditioned pain modulation). Il peut être utilisé en thérapeutique pour inhiber une douleur grâce à une stimulation nociceptive supportable. C'est par exemple le cas d'un traitement de la crise de migraine désigné sous le terme de remote electrical neuromodulation (stimulation douloureuse brachiale permettant de réduire la douleur migraineuse).
- La douleur est indispensable et contribue à la protection de l'intégrité de l'organisme.
- Les neurones de 1er ordre sont de petites fibres, peu ou pas myélinisées (A,δ et C). Leurs terminaisons nerveuses libres répondent à différents stimuli.
- La voie néospinothalamique correspond à la voie sensoridiscriminative qui véhicule les informations nociceptives.
Voir QRM chapitre 32.
Pour en savoir plus
- Basbaum AI, Bautista DM, Scherrer G, Julius D. Cellular and molecular mechanisms of pain. Cell 2009 ;139:267–84.