Laurent Tatu et Fabrice Vuillier
Anatomie du cerveau
Laurent Tatu et Fabrice Vuillier
Le cerveau appartient à l'encéphale avec le tronc cérébral et le cervelet. Il comporte une partie centrale, le diencéphale, qui permet la communication avec le tronc cérébral, et deux hémisphères qui forment le télencéphale.
La substance grise du cerveau qui renferme les neurones est répartie en substance grise profonde (essentiellement noyau lentiforme, noyau caudé et thalamus) et en substance grise périphérique correspondant au cortex cérébral.
La substance blanche qui représente les fibres nerveuses se place dans le centre ovale des hémisphères, dans les régions de concentrations de fibres appelées capsules (capsules interne et externe) et dans les commissures qui unissent morphologiquement et fonctionnellement les deux hémisphères (corps calleux, fornix, commissures antérieure et postérieure).
Le liquide cérébrospinal se répartit dans le 3e ventricule, qui creuse le diencéphale, et dans les ventricules latéraux présents dans chaque hémisphère (1er et 2e ventricules).
Anatomie du cortex cérébral (figures 1.1 à 1.3)
La substance grise périphérique du cerveau forme le cortex cérébral à la surface des hémisphères cérébraux. Le cortex humain a un aspect plissé définissant la gyrencéphalie qui cache les deux tiers du cortex cérébral en profondeur des plis présents à la surface du cerveau.
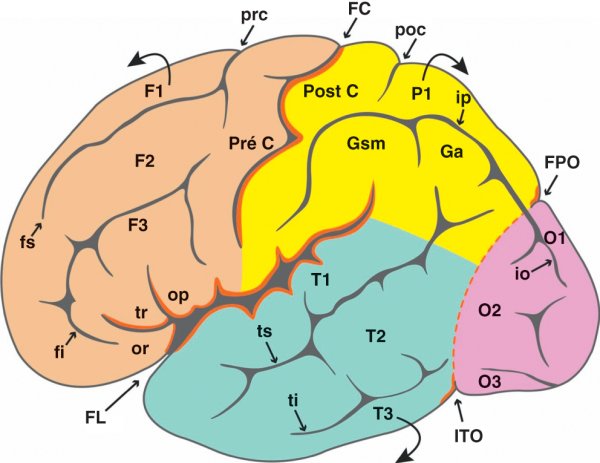
Figure 1.1
Vue latérale de l'hémisphère cérébral.
FC : fissure centrale ; FL : fissure latérale ; FPO : fissure pariéto-occipitale ; ITO : incisure temporo-occipitale. Lobe frontal (beige) – F1 : gyrus frontal supérieur ; F2 : gyrus frontal moyen ; F3 : gyrus frontal inférieur ; fi : sillon frontal inférieur ; fs : sillon frontal supérieur ; op : pars operculaire ; or : pars orbitaire ; prc : sillon précentral ; Pré C : gyrus précentral ; tr : pars triangulaire. Lobe temporal (vert) – T1 : gyrus temporal supérieur ; T2 : gyrus temporal moyen ; T3 : gyrus temporal inférieur ; ti : sillon temporal inférieur ; ts : sillon temporal supérieur. Lobe pariétal (jaune) – Ga : gyrus angulaire ; Gsm : gyrus supramarginal ; ip : sillon intrapariétal ; P1 : gyrus pariétal supérieur ; poc : sillon post-central ; Post C : gyrus post-central. Lobe occipital (rose) – io : sillon intra-occipital ; O1 : gyrus occipital supérieur ; O2 : gyrus occipital moyen ; O3 : gyrus occipital inférieur.
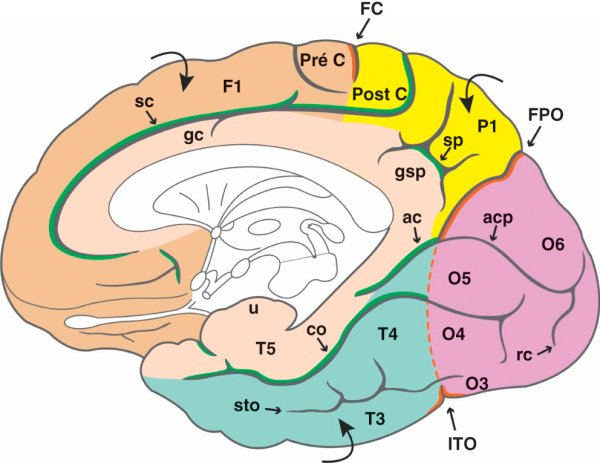
Figure 1.2
Vue médiale de l'hémisphère cérébral.
FC : fissure centrale ; FPO : fissure pariéto-occipitale ; ITO : incisure temporo-occipitale. Lobe limbique (chair) – ac : sillon antécalcarin ; co : sillon collatéral ; gc : gyrus cingulaire ; gsp : gyrus subpariétal ; sc : sillon cingulaire ; sp : sillon subpariétal ; T5 : gyrus parahippocampique ; u : uncus. Lobe frontal (beige) – F1 : gyrus frontal supérieur ; Pré C : gyrus précentral. Lobe pariétal (jaune) – P1 : gyrus pariétal supérieur ; Post C : gyrus post-central. Lobe temporal (vert) – sto : sillon temporo-occipital ; T3 : gyrus temporal inférieur ; T4 : gyrus fusiforme. Lobe occipital (rose) – acp : sillon calcarin proprement dit ; O3 : gyrus occipital inférieur ; O4 : 4e gyrus occipital ; O5 : gyrus lingual ; O6 : cunéus ; rc : sillon rétrocalcarin.
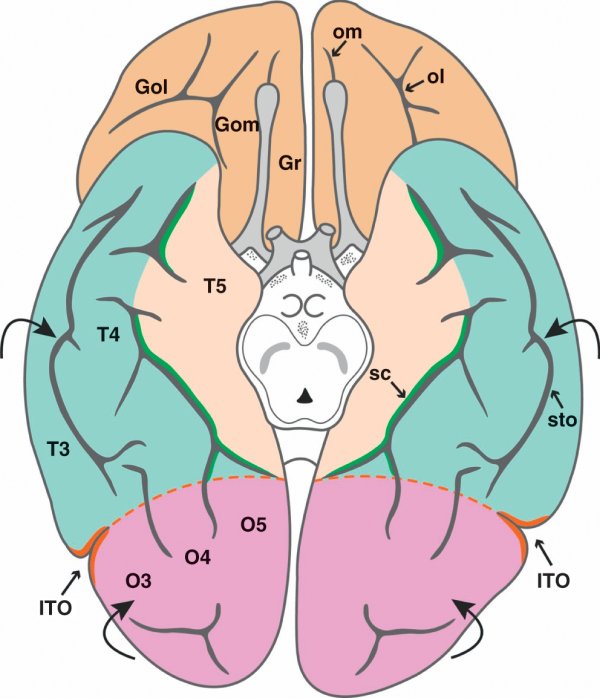
Figure 1.3
Vue inférieure de l'hémisphère cérébral.
Gol : gyrus orbitaire latéral ; Gom : gyrus orbitaire médial ; Gr : gyrus rectus ; ITO : incisure temporo-occipitale ; O3 : gyrus occipital inférieur ; O4 : 4e gyrus occipital ; O5 : gyrus lingual ; ol : sillon orbitaire latéral ; om : sillon orbitaire médial ; sc : sillon cingulaire ; sto : sillon temporo-occipital ; T3 : gyrus temporal inférieur ; T4 : gyrus fusiforme ; T5 : gyrus parahippocampique.
Lobe frontal
Sur la face latérale du lobe frontal, se trouvent le sillon frontal supérieur et le sillon frontal inférieur. Ces deux sillons à disposition horizontale interceptent à angle droit un sillon vertical, le sillon précentral. Le gyrus frontal supérieur (F1) se place au-dessus du sillon frontal supérieur. Le gyrus frontal moyen (F2) est situé entre les sillons frontaux supérieur et inférieur. Le gyrus frontal inférieur (F3) est placé en dessous du sillon frontal inférieur. Il est subdivisé par la présence des deux rameaux de la fissure latérale en une partie operculaire, en arrière du rameau vertical, une partie triangulaire entre les rameaux vertical et horizontal, et une partie orbitaire en avant du rameau horizontal. Le gyrus précentral est à disposition verticale entre le sillon précentral et la fissure centrale.
La face médiale du lobe frontal est marquée par la présence du gyrus frontal supérieur (F1) qui se poursuit depuis la convexité cérébrale. La fissure centrale, les gyrus précentral et post-central débordent également sur cette face médiale en formant le lobule paracentral qui correspond aux centres moteurs et sensitifs du membre inférieur controlatéral.
La face inférieure, également appelée face orbitaire, comporte le sillon orbitaire médial, à disposition rectiligne, également appelé sillon olfactif car il accueille le bulbe et le tractus olfactifs, et le sillon orbitaire latéral qui prend habituellement la forme de la lettre H. Le gyrus rectus est compris entre la fissure interhémisphérique et le sillon orbitaire médial. Le gyrus orbitaire médial est compris entre le sillon orbitaire médial et le sillon orbitaire latéral. Le gyrus orbitaire latéral est compris entre le sillon orbitaire latéral et la convexité cérébrale.
Lobe temporal
Il est situé en dessous de la fissure latérale. Sa limite postérieure est matérialisée par une ligne théorique qui unit la fissure pariéto-occipitale à l'incisure temporo-occipitale.
Sur la face latérale apparaissent le sillon temporal supérieur, parallèle à la fissure latérale, et le sillon temporal inférieur. Le gyrus temporal supérieur (T1) se place entre la fissure latérale et le sillon temporal supérieur. Le gyrus temporal moyen (T2) est délimité par les sillons temporaux supérieur et inférieur. Le gyrus temporal inférieur (T3) est situé en dessous du sillon temporal inférieur.
Sur la face médiale se placent le sillon temporo-occipital, qui se prolonge dans le lobe occipital, et le sillon collatéral. Le sillon temporo-occipital sépare le gyrus temporal inférieur (T3) du gyrus fusiforme (T4), lui-même séparé du gyrus parahippocampique (T5), qui appartient au lobe limbique, par le sillon collatéral. La partie antérieure du gyrus parahippocampique est marquée par la présence de l'uncus, un lobule cortical recourbé en arrière et qui appartient au lobe limbique.
Lobe pariétal
Il est situé en arrière du lobe frontal, dont il est séparé par la fissure centrale, et en avant du lobe occipital, dont il est séparé par la fissure pariéto-occipitale prolongée par une ligne théorique se dirigeant vers l'incisure temporo-occipitale. Cette région postérieure aux contours flous correspond au carrefour temporopariéto-occipital.
Le sillon intrapariétal traverse la face latérale comme un arc de cercle pour se terminer en sillon intra-occipital dans le lobe occipital. La partie initiale ascendante du sillon intrapariétal correspond au sillon post-central inférieur qui constitue, avec son homologue le sillon post-central supérieur, le sillon post-central. Le gyrus post-central est compris entre le sillon post-central et la fissure centrale. Au-dessus du sillon intrapariétal se place le gyrus pariétal supérieur (P1). En dessous du sillon intrapariétal se situe le gyrus pariétal inférieur (P2), qui est subdivisé en gyrus supramarginal à l'extrémité de la fissure centrale et en gyrus angulaire à l'extrémité du sillon temporal supérieur.
La face médiale du lobe pariétal est marquée par la présence du sillon subpariétal qui sépare le gyrus pariétal supérieur (P1) du gyrus subpariétal appartenant au lobe limbique. La partie la plus postérieure du lobule paracentral est également visible.
Lobe occipital
Il est situé à la partie postérieure et inférieure de l'hémisphère cérébral en arrière des lobes pariétal et temporal dont il est séparé par une limite théorique allant de la fissure pariéto-occipitale, bien visible sur la face médiale de l'hémisphère, à l'incisure temporo-occipitale. L'organisation sulcogyrale du lobe occipital est complexe.
Le sillon majeur de la face médiale du lobe occipital est le sillon calcarin qui peut être divisé en trois parties : le sillon antécalcarin, qui appartient à la fissure limbique, le sillon calcarin proprement dit, d'aspect sinueux, et le sillon rétrocalcarin à disposition verticale. Sur cette face médiale, le gyrus occipital inférieur (O3) se place dans la continuité du gyrus temporal inférieur (T3) ; le quatrième gyrus occipital (O4) prolonge le gyrus fusiforme (T4) ; le gyrus lingual (O5) prolonge le gyrus parahippocampique (T5). Le seul gyrus occipital bien délimité sur la face médiale est le cunéus (O6) qui s'intercale entre la fissure pariéto-occipitale et le sillon calcarin
Sur la face latérale, se trouve le sillon intra-occipital, terminaison du sillon intrapariétal. Le gyrus occipital supérieur (O1) se place dans le prolongement du gyrus pariétal supérieur (P1) et les gyrus occipitaux moyen (O2) et inférieur (O3) sont situés dans la continuité des gyrus temporaux correspondants T2 et T3.
Lobe limbique et hippocampe (figures 1.2 et 1.4)
La face médiale de l'hémisphère cérébral porte le lobe limbique situé autour du diencéphale et regroupant l'achéocortex. Il constitue une partie des lobes frontal, pariétal et temporal. Le lobe limbique est séparé du reste du cortex par la fissure limbique constituée d'une succession de sillons parmi lesquels :
- le sillon cingulaire qui suit la courbe du corps calleux puis se redresse pour atteindre le bord supérieur de l'hémisphère ;
- le sillon subpariétal en forme de H ;
- le sillon antécalcarin qui correspond à la partie antérieure du sillon calcarin du lobe occipital ;
- le sillon collatéral du lobe temporal.
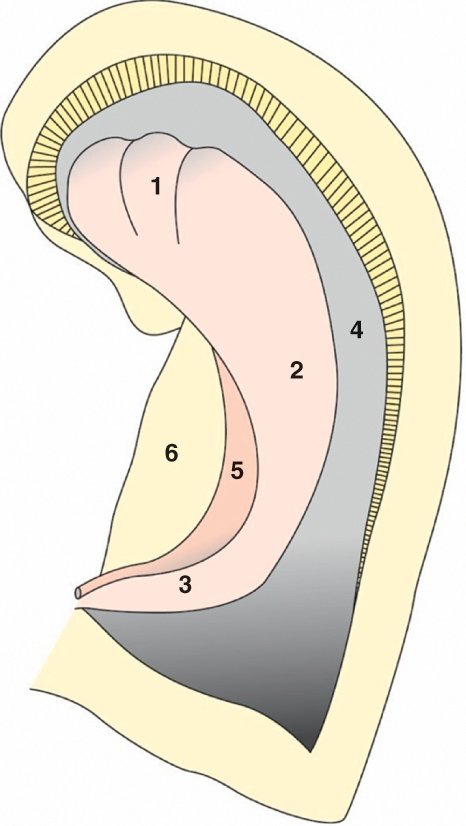 |
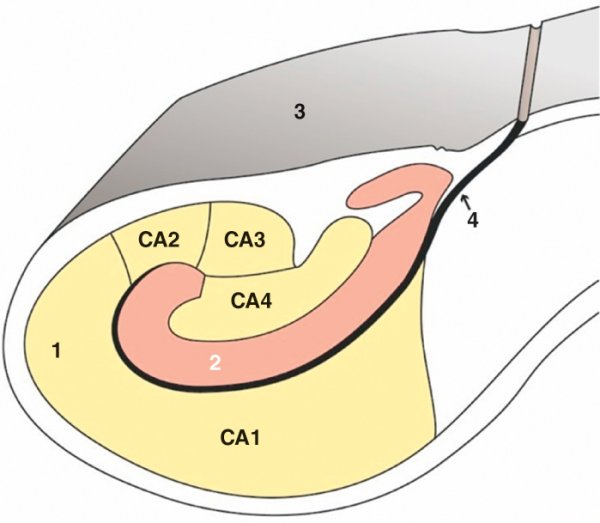 |
Figure 1.4
Hippocampe.
A. Vue supérieure endoventriculaire de l'hippocampe droit. 1. Tête de l'hippocampe. 2. Corps de l'hippocampe. 3. Queue de l'hippocampe. 4. Corne ventriculaire temporale ouverte. 5. Fimbria. 6. Subiculum. B. Coupe frontale de l'hippocampe. 1. Corne d'Ammon (champs CA1, CA2, CA3, CA4). 2. Gyrus dentatus. 3. Subiculum. 4. Sillon hippocampique.
Le lobe limbique, situé en dedans de cette fissure, est divisé en gyrus subcalleux et subpariétal en dessous des sillons du même nom, et en gyrus parahippocampique (T5) qui recouvre l'hippocampe, également constitué d'archéocortex, et le noyau amygdalien.
L'hippocampe est formé de deux lames d'archéocortex, le gyrus dentatus et la corne d'Ammon, enroulées l'une dans l'autre. Il est situé dans la corne temporale du ventricule latéral et peut être divisé en trois parties : tête, corps et queue. La terminaison effilée des piliers du fornix, formant la fimbria, vient s'accoler à l'hippocampe. L'hippocampe est séparé d'une zone corticale temporale aplatie, appelée subiculum, par le sillon de l'hippocampe. La partie antérieure de l'hippocampe est au contact d'une autre structure importante : le noyau amygdalien.
Lobe de l'insula (figure 1.5)
L'insula est la seule partie du cortex cérébral qui n'est pas visible à la surface du cerveau. Enfouie au fond de la fissure latérale, elle est recouverte par d'autres zones du cortex formant un opercule à sa surface. Les lobes frontal, pariétal et temporal participent à la constitution de cet opercule insulaire. Dans sa partie supérieure, l'insula est recouverte par l'opercule frontopariétal, qui est constitué, d'avant en arrière, par les pars triangularis et opercularis du gyrus frontal inférieur (F3), par le gyrus subcentral qui coiffe l'extrémité inférieure de la fissure centrale, par la partie inférieure du gyrus post-central et par le gyrus supramarginal qui coiffe l'extrémité de la fissure latérale. Dans sa partie inférieure, le cortex insulaire est recouvert par l'opercule temporal, constitué par la face supérieure du gyrus temporal supérieur (T1) qui porte les gyrus temporaux transverses (Heschl). Dans sa partie antérieure, l'insula est recouverte par l'opercule fronto-orbitaire, qui comprend les gyrus de la face orbitaire du lobe frontal.
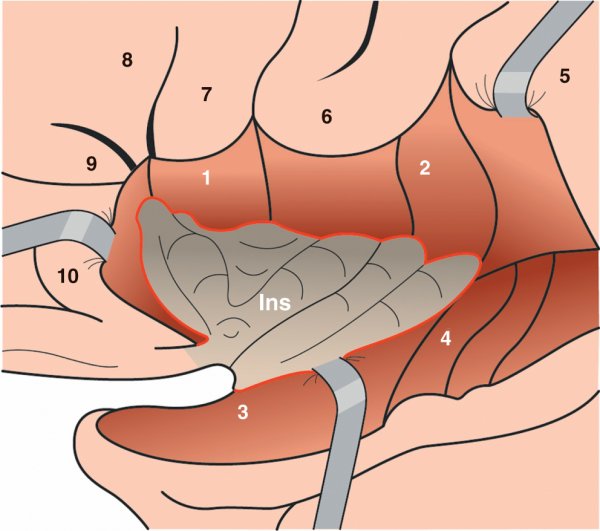
Figure 1.5
Insula (Ins).
1. Opercule frontal. 2. Opercule pariétal. 3. Opercule temporal. 4. Gyrus temporaux transverses. 5. Gyrus supramarginal. 6. Gyrus subcentral. 7. Pars operculaire du gyrus orbitaire inférieur. 8. Pars triangulaire du gyrus orbitaire inférieur. 9. Pars orbitaire du gyrus orbitaire inférieur. 10. Gyrus orbitaires.
Configuration intérieure du cerveau
Substance grise profonde (figures 1.6 et 1.7)
Elle est regroupée en noyaux diencéphaliques (essentiellement le thalamus) et télencéphaliques (essentiellement noyaux lentiforme et caudé).
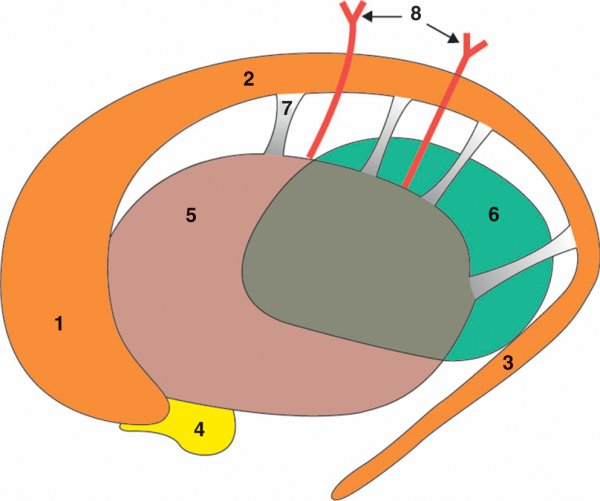
Figure 1.6
Noyaux profonds de substance grise.
1. Tête du noyau caudé. 2. Corps du noyau caudé. 3. Queue du noyau caudé. 4. Nucleus accumbens (septi). 5. Noyau lentiforme. 6. Thalamus. 7. Pont putaminocaudé. 8. Faisceau corticospinal.
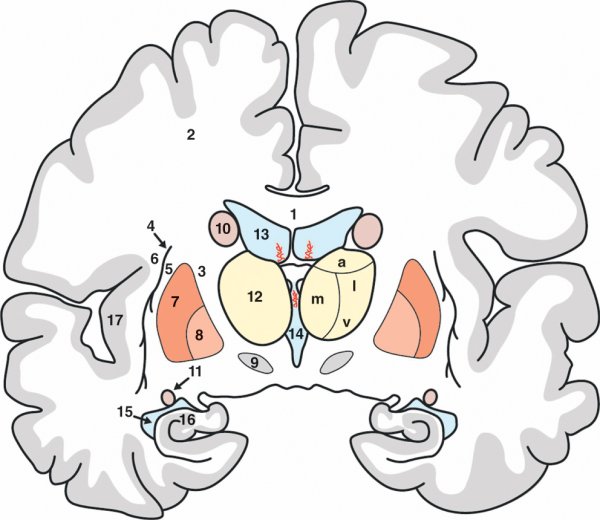
Figure 1.7
Coupe frontale du cerveau.
1. Corps calleux. 2. Centre ovale. 3. Capsule interne. 4. Claustrum. 5. Capsule externe. 6. Capsule extrême. 7. Putamen. 8. Globus pallidus. 9. Noyau subthalamique. 10. Corps du noyau caudé. 11. Queue du noyau caudé. 12. Thalamus. Groupes nucléaires antérieur (a), médial (m), latéral (l) et ventral (v). 13. Corps du ventricule latéral. 14. Troisième ventricule. 15. Corne temporale du ventricule latéral. 16. Hippocampe. 17. Insula
Le noyau caudé est une structure arciforme qui s'enroule autour du noyau lentiforme. Il possède trois segments : la tête collée en avant au noyau lentiforme, le corps relié au noyau lentiforme par des ponts putaminocaudés, et la queue qui part de manière effilée en direction du lobe temporal de l'hémisphère. À la jonction de la tête du noyau caudé et du noyau lentiforme se place le nucleus accumbens (septi).
Le noyau lentiforme prend un aspect triangulaire à la coupe avec une partie périphérique appelée putamen (la coquille en latin) et une partie centrale plus pâle dénommée globus pallidus qui est divisée sur un plan fonctionnel en globus pallidus externe et globus pallidus interne.
En position plus médiale par rapport au complexe noyau caudé – noyau lentiforme, se place le thalamus, noyau diencéphalique de substance grise. Il constitue le relais obligatoire des voies sensitives et sensorielles. Les corps géniculés qui lui sont annexés forment des relais de la vision (corps géniculé latéral) et de l'audition (corps géniculé médial). Le thalamus est lui-même subdivisé en sous-groupes nucléaires antérieur, médial, ventral et latéral. Le groupe ventral est la zone de relais des voies cérébelleuses et sensitives.
Le thalamus surplombe lui-même un autre noyau faisant la jonction avec le mésencéphale, le noyau subthalamique qui appartient aux ganglions de la base et intervient dans la régulation de la motricité.
Les commissures cérébrales correspondent à des faisceaux de substance blanche qui assurent la transmission des informations entre les deux hémisphères. La plus volumineuse d'entre elles correspond au corps calleux qui est divisé en plusieurs parties d'avant en arrière : le bec, le genou, le corps et le splenium, la portion la plus dilatée et fonctionnellement la plus importante.
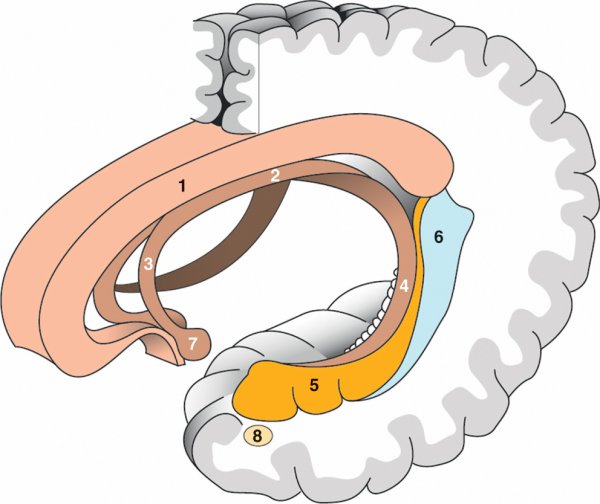
Figure 1.8
Commissures cérébrales.
1. Corps calleux. 2. Corps du fornix. 3. Colonne du fornix. 4. Pilier du fornix se prolongeant par la fimbria. 5. Hippocampe. 6. Corne temporale du ventricule latéral. 7. Corps mamillaire. 8. Noyau amygdalien.
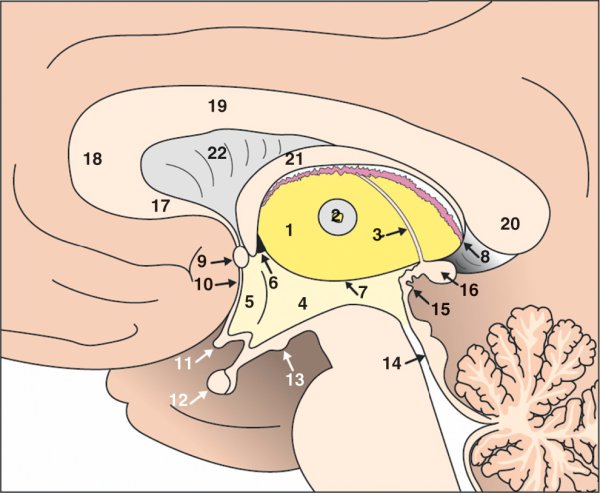
Figure 1.9
Coupe sagittale du diencéphale.
1. Thalamus. 2. Adhésion interthalamique. 3. Habenula. 4. Hypothalamus. 5. Aire préoptique. 6. Foramen interventriculaire. 7. Sillon hypothalamique. 8. Plexus choroïdes. 9. Commissure antérieure. 10. Lame terminale. 11. Chiasma optique. 12. Hypophyse. 13. Corps mamillaire. 14. Aqueduc du mésencéphale. 15. Commissure postérieure. 16. Glande pinéale. 17. Bec du corps calleux. 18. Genou du corps calleux. 19. Corps du corps calleux. 20. Splénium du corps calleux. 21. Fornix. 22. Septum lucidum.
Une autre commissure est constituée par le fornix qui se place en dessous du corps calleux. À partir d'un corps aplati rattaché au corps calleux par une membrane translucide, le septum lucidum, le fornix émet des prolongements vers les deux hémisphères. Les prolongements antérieurs, appelés colonnes, partent en direction des corps mamillaires, et les prolongements postérieurs, les piliers, viennent s'accoler à l'hippocampe en formant la fimbria. Le rôle fonctionnel du fornix est majeur pour le lobe limbique.
Deux autres commissures, de forme plus tubulaire, correspondent aux commissures antérieure et postérieure. L'alignement horizontal correct de ces deux commissures sur une coupe sagittale sert, en imagerie, à la définition du plan de référence bicommissural.
Substance blanche intrahémisphérique (cf. figure 1.7)
Elle possède des zones de haute concentration de fibres axonales appelées capsules. La capsule interne se glisse entre le thalamus en dedans et le noyau caudé et le noyau lentiforme en dehors. Les fibres motrices et sensitives y transitent. Entre le putamen et l'insula, une lame de substance grise appelée claustrum délimite les capsules externe et extrême. La vaste nappe de substance blanche qui surplombe l'étage des capsules correspond au centre ovale.
enroulement du cerveau. Le corps du ventricule émet des prolongements appelés cornes dans les lobes frontal, occipital et temporal.
Le ventricule diencéphalique correspond au 3e ventricule. Cette cavité liquidienne est limitée en avant par la commissure antérieure, la lame terminale, une fine lame de substance blanche, et le chiasma optique. Sa limite inférieure ou plancher correspond au losange optopédonculaire avec la présence de la tige de l'hypophyse et des corps mamillaires. Sa paroi postérieure est faite de la commissure postérieure surplombée par la glande pinéale et sa limite supérieure est marquée par les deux habenulas et les plexus choroïdes. Les deux parois latérales du 3e ventricule sont symétriques et représentées, dans la partie haute, par les thalamus fréquemment reliés par l'adhésion interthalamique et, dans la partie basse, par l'hypothalamus. L'hypothalamus est séparé du thalamus par le sillon hypothalamique et il présente dans sa partie antérieure l'aire préoptique.
Le 3e ventricule communique avec chacun des ventricules latéraux par le foramen interventriculaire et avec le 4e ventricule par l'aqueduc du mésencéphale.
Anatomie du tronc cérébral et du cervelet
Laurent Tatu et Fabrice Vuillier
Anatomie du tronc cérébral (figures 1.10 et 1.11)
Le tronc cérébral est situé avec le cervelet dans la fosse crânienne postérieure. Il fait directement suite à la moelle spinale et se prolonge par le diencéphale. Le 4e ventricule s'intercale entre le tronc cérébral et le cervelet, structures reliées par les pédoncules cérébelleux.
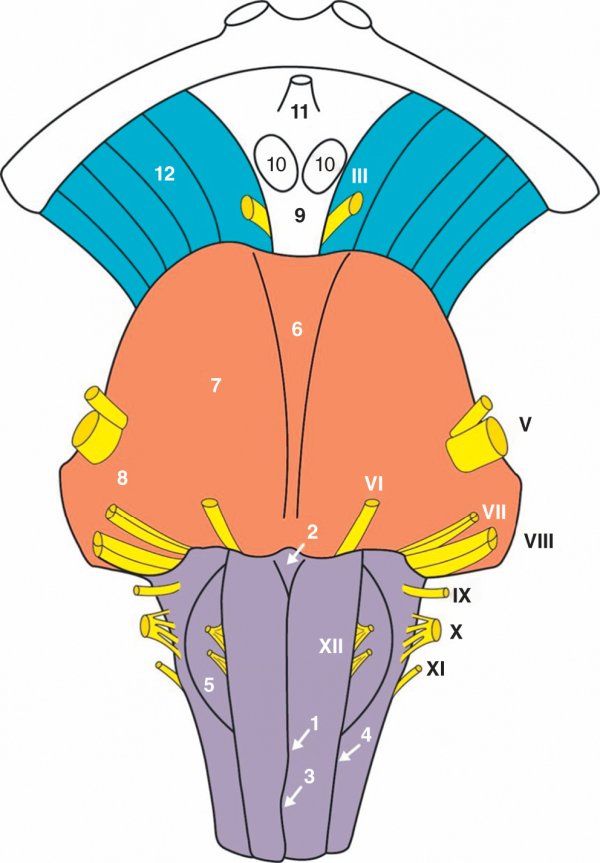
Figure 1.10
Vue ventrale du tronc cérébral.
1. Sillon médian du bulbe. 2. Foramen cæcum. 3. Décussation motrice. 4. Sillon latéral du bulbe (préolivaire). 5. Olive bulbaire. 6. Sillon basilaire. 7. Pyramide du pont. 8. Pédoncule cérébelleux moyen. 9. Losange optopédonculaire. 10. Corps mamillaire. 11. Tige de l'hypophyse. 12. Pédoncule cérébral.
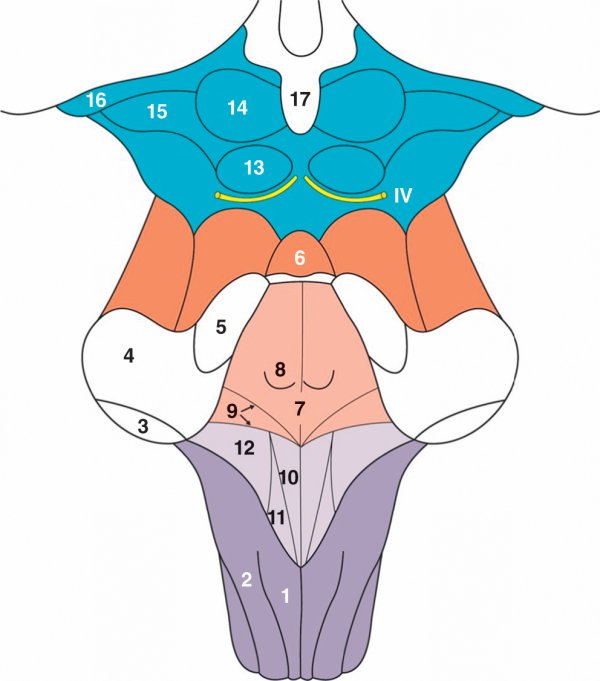
Figure 1.11
Vue dorsale du tronc cérébral.
1. Tubercule gracile. 2. Tubercule cunéiforme. 3. Pédoncule cérébelleux inférieur. 4. Pédoncule cérébelleux moyen. 5. Pédoncule cérébelleux supérieur. 6. Voile médullaire supérieur. 7. Plancher du 4e ventricule. 8. Colliculus facial. 9. Stries médullaires. 10. Trigone de l'hypoglosse. 11. Trigone du vague. 12. Aire vestibulaire. 13. Colliculus inférieur. 14. Colliculus supérieur. 15. Corps géniculé médial. 16. Corps géniculé latéral. 17. Glande épiphyse.
Sur un plan morphologique, le tronc cérébral est segmenté en trois parties dans le sens craniocaudal : le mésencéphale, le pont et le bulbe (moelle allongée). Au niveau bulbaire et pontique, le plancher du 4e ventricule constitue la face dorsale du tronc cérébral.
Bulbe (moelle allongée)
Il fait directement suite à la moelle spinale cervicale. Il comporte deux étages : une partie inférieure fermée, qui présente la même organisation que la moelle spinale cervicale, et un étage supérieur ouvert par la présence du 4e ventricule.
Sa partie ventrale est marquée par la présence des pyramides bulbaires séparées par le sillon médian du bulbe qui se dilate dans sa partie haute pour former un point d'entrée vasculaire appelé foramen cæcum. Sur la partie basse de ce sillon apparaît la décussation motrice. Les olives inférieures (olives bulbaires) apparaissent sur la partie latérale en arrière du sillon préolivaire.
La face dorsale de la partie inférieure du bulbe permet de voir la disposition des faisceaux postérieurs sensitifs gracile (de Goll) et cunéiforme (de Burdach) se dilatant dans leur partie haute en tubercules du même nom. Dans la partie supérieure, la face dorsale disparaît pour laisser apparaître le plancher du 4e ventricule encadré par les pédoncules cérébelleux inférieurs. Cette partie bulbaire du plancher du 4e ventricule est séparée de la partie pontique par les stries médullaires. Les trigones du nerf hypoglosse et du nerf vague ainsi que l'aire vestibulaire font saillie dans la partie bulbaire.
Pont
Il constitue une saillie transversale qui sépare le bulbe du mésencéphale. Il se prolonge latéralement par les pédoncules cérébelleux moyens. Le pont est marqué sur la ligne médiane par la présence du sillon basilaire, une dépression qui livre passage à l'artère du même nom. Il se prolonge latéralement par les pyramides du pont qui se dirigent vers les pédoncules cérébelleux moyens.
La face dorsale du pont constitue le plancher du 4e ventricule où les saillies du colliculus facial (relief des noyaux des nerfs abducens et facial) sont visibles. Dans la partie haute du pont, les pédoncules cérébelleux supérieurs sont unis par le voile médullaire supérieur.
Mésencéphale
Sur la face ventrale du mésencéphale, les éléments les plus saillants correspondent aux pédoncules cérébraux qui s'infiltrent dans le diencéphale. Ils sont séparés par le losange optopédonculaire, délimité en avant par le chiasma optique, et qui comporte les corps mamillaires et la tige de l'hypophyse.
Sur la face dorsale, les colliculus supérieur et inférieur sont reliés respectivement aux corps géniculés latéral et médial qui se collent au thalamus. La face dorsale du mésencéphale est surplombée par la glande épiphyse.
Origine apparente des nerfs crâniens
Chacun des dix nerfs crâniens vrais possède une origine apparente à la surface du tronc cérébral. Seul le nerf trochléaire (IV) émerge de la face dorsale du tronc cérébral au niveau des colliculus inférieurs. Les neuf autres émergent de la face ventrale :
- le nerf oculomoteur (III) dans la fosse interpédonculaire ;
- le nerf trijumeau (V) et ses deux racines, motrice et sensitive, sur la face latérale du pont ;
- le nerf abducens (VI) dans le sillon bulbopontique ;
- les nerfs facial (VII), intermédiaire (VIIbis) et cochléovestibulaire (VIII) dans l'angle pontocérébelleux ;
- les nerfs glossopharyngien (IX), vague (X) et accessoire (XI) dans la fossette rétro-olivaire ;
- le nerf hypoglosse (XII) dans le sillon préolivaire.
Configuration intérieure du tronc cérébral (figures 1.12 à 1.14)
Le tronc cérébral, comme le reste du système nerveux central, est formé de substance blanche, de substance grise et de liquide cérébrospinal contenu dans le 4e ventricule.
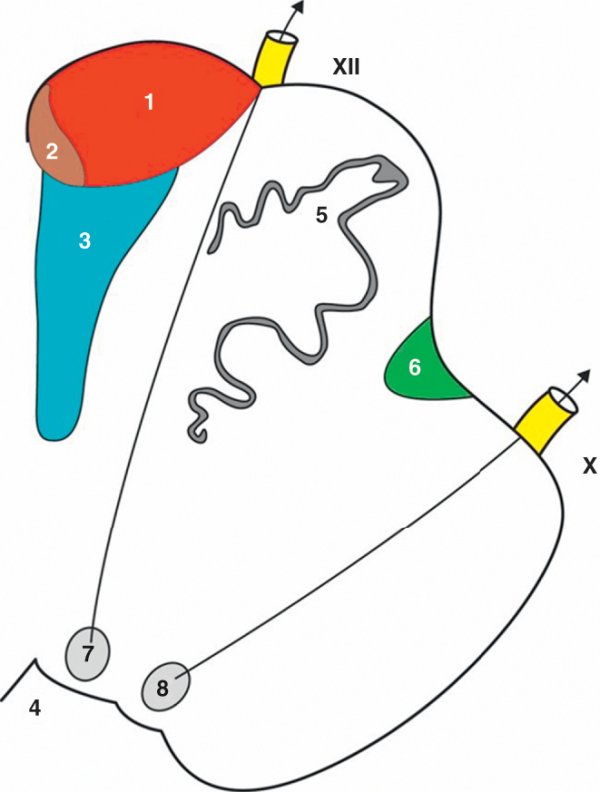
Figure 1.12
Coupe horizontale du bulbe ouvert.
1. Faisceau corticospinal. 2. Faisceau corticonucléaire. 3. Lemniscus médial. 4. 4e ventricule. 5. Olive bulbaire. 6. Faisceau spinothalamique. 7. Noyau du nerf hypoglosse. 8. Noyau dorsal moteur du nerf vague. X : origine apparente du nerf vague ; XII : origine apparente du nerf hypoglosse.
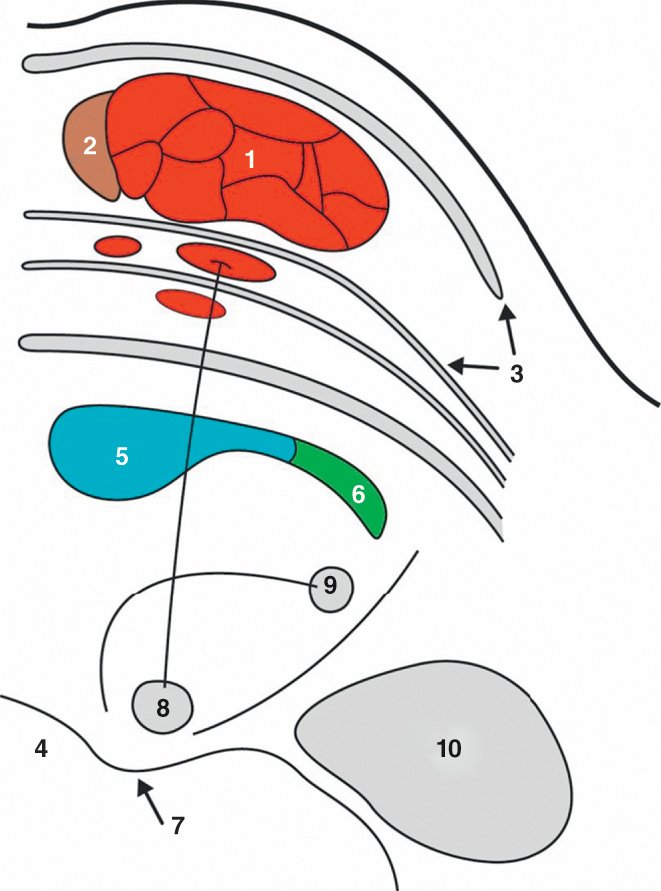
Figure 1.13
Coupe horizontale du pont (passant par le colliculus facial).
1. Faisceau corticospinal. 2. Faisceau corticonucléaire. 3. Noyaux du pont. 4. 4e ventricule. 5. Lemniscus médial. 6. Faisceau spinothalamique. 7. Colliculus facial. 8. Noyau du nerf abducens. 9. Noyau du nerf facial. 10. Noyaux cochléovestibulaires.
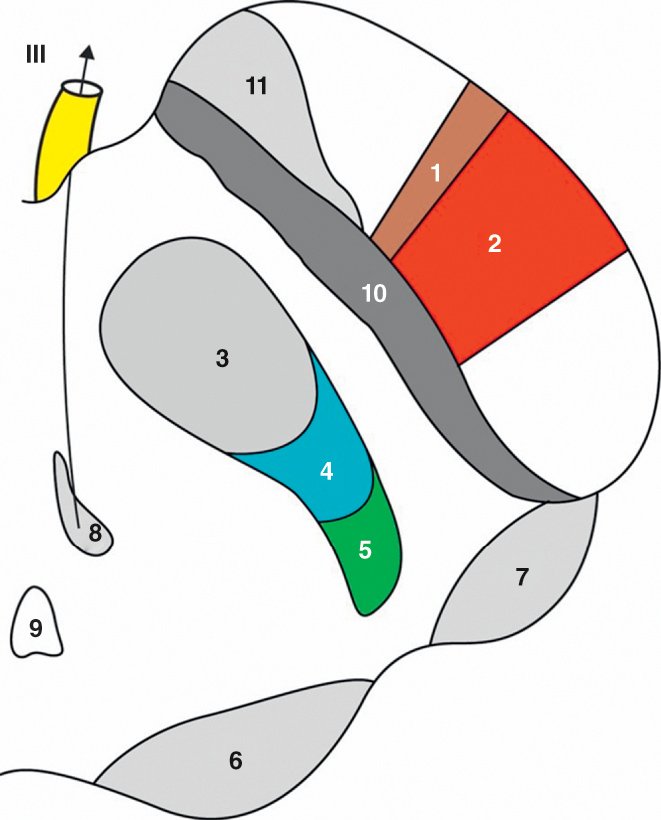
Figure 1.14
Coupe horizontale du mésencéphale (passant par le colliculus supérieur).
1. Faisceau corticonucléaire. 2. Faisceau corticospinal. 3. Noyau rouge. 4. Lemniscus médial. 5. Faisceau spinothalamique. 6. Colliculus supérieur. 7. Corps géniculé médial. 8. Noyau du nerf moteur oculaire. 9. Aqueduc du mésencéphale. 10. Substance noire compacte. 11. Substance noire réticulaire.
La substance grise du tronc cérébral est répartie en noyaux des nerfs crâniens, substance réticulée et noyaux propres du tronc cérébral. Les noyaux des nerfs crâniens sont organisés en colonnes plaquées en regard du plancher du 4e ventricule. Certains d'entre eux peuvent être localisés par leurs reliefs de surface comme le colliculus facial (repère des noyaux des nerfs abducens et facial), le trigone de l'hypoglosse ou encore le trigone du vague.
Les noyaux propres du tronc cérébral sont des relais importants sur les voies de la motricité et de la sensibilité. Les principaux de ces noyaux sont :
- le noyau rouge, la substance noire et les noyaux des colliculus au niveau mésencéphalique ;
- les noyaux du pont au niveau pontique ;
- les noyaux gracile et cunéiforme et l'olive inférieure (bulbaire) au niveau bulbaire.
La formation réticulaire est organisée en groupes nucléaires en position centrale dans le tronc cérébral, elle échappe à la visualisation macroscopique.
La substance blanche du tronc cérébral est formée des faisceaux de passage descendants moteurs et ascendants sensitifs, des faisceaux sensoriels et des faisceaux propres du tronc cérébral, comme le faisceau longitudinal médial, et de la partie intra-axiale des fibres des nerfs crâniens.
Le faisceau corticospinal occupe le tiers médian du pédoncule cérébral. Il est divisé en une multitude de fascicules lors de la traversée du pont et il se reconstitue pour traverser le bulbe dans la pyramide bulbaire. Il décusse à la partie inférieure du bulbe et seule une petite partie des fibres forme le faisceau corticospinal direct. Le faisceau corticonucléaire qui envoie ses fibres, après décussation, vers les noyaux des nerfs crâniens est situé médialement par rapport au précédent. Les voies sensitives sont réparties dans le lemniscus médial pour la sensibilité épicritique et le faisceau spinothalamique pour la sensibilité protopathique. Le lemniscus médial est en situation médiale et verticale au niveau bulbaire alors que le faisceau spinothalamique est situé plus latéralement. Dans le pont, le lemniscus médial s'horizontalise et se colle au faisceau spinothalamique. Dans le pédoncule cérébral, les deux voies sensitives viennent s'accoler au noyau rouge.
Anatomie du cervelet (figures 1.15 et 1.16)
Le cervelet, logé dans la fosse crânienne postérieure, est séparé du tronc cérébral en avant de lui par le 4e ventricule. Avec le bulbe et le pont, il forme le rhombencéphale. Par sa face antérieure, il forme donc le toit de la cavité du 4e ventricule et communique avec le tronc cérébral par trois paires de pédoncules cérébelleux : inférieurs, moyens et supérieurs. Les pédoncules cérébelleux véhiculent les faisceaux afférents et efférents qui unissent le cervelet au reste du système nerveux central. Sa face supérieure est au contact de la structure dure-mérienne appelée tente du cervelet qui ferme en haut la fosse crânienne postérieure. Sa face inférieure est au contact de l'os occipital. La fissure horizontale, habituellement bien marquée, fait le tour des hémisphères dans le plan horizontal.
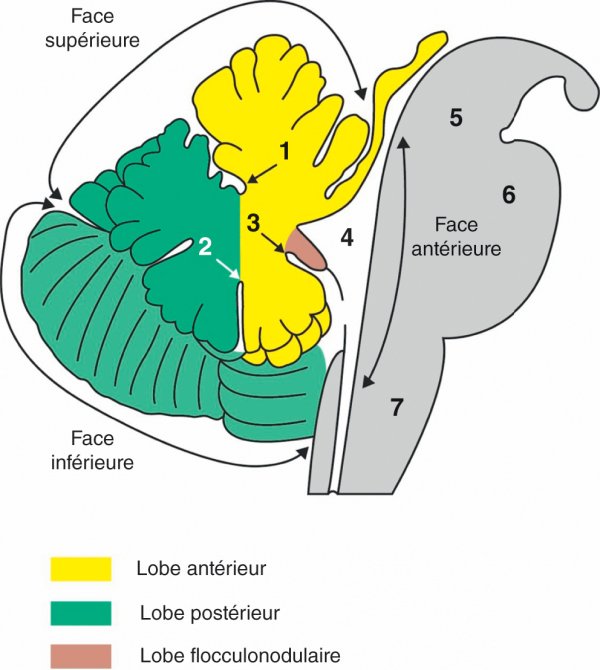
Figure 1.15
Coupe sagittale médiane du tronc cérébral et du cervelet.
1. Fissure primaire. 2. Fissure secondaire. 3. Fissure postérolatérale. 4. 4e ventricule. 5. Mésencéphale. 6. Pont. 7. Bulbe (moelle allongée).
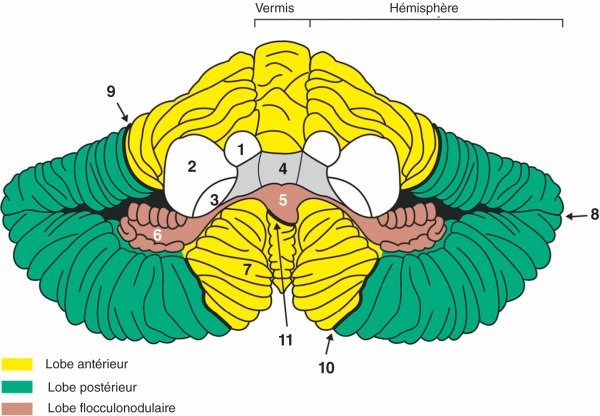
Figure 1.16
Vue ventrale du cervelet.
1. Pédoncule cérébelleux supérieur. 2. Pédoncule cérébelleux moyen. 3. Pédoncule cérébelleux inférieur. 4. 4e ventricule. 5. Nodule. 6. Floccule. 7. Tonsille. 8. Fissure horizontale. 9. Fissure primaire. 10. Fissure secondaire. 11. Fissure postérolatérale.
Le cervelet est divisé en une partie médiane étroite, le vermis, et deux parties latérales développées, les hémisphères cérébelleux. Il est divisé en trois lobes : le lobe antérieur séparé du lobe postérieur par la fissure primaire et la fissure secondaire, et le lobe flocculonodulaire séparé du lobe postérieur par la fissure postérolatérale. Le lobe flocculonodulaire est constitué du nodule qui appartient au vermis prolongé latéralement par les floccules qui se placent sur la face ventrale de chaque hémisphère cérébelleux.
À l'image du cerveau, le cervelet comporte de la substance blanche, de la substance grise périphérique, le cortex, et de la substance grise centrale. Le cortex cérébelleux est plissé et peut être divisé en une série de lobules. Parmi ceux-ci, les tonsilles occupent une place stratégique en surplombant le foramen magnum au contact proche du bulbe. Ces structures sont responsables de l'engagement cérébelleux en cas d'hyperpression dans la fosse crânienne postérieure.
La substance grise centrale est organisée en trois paires de noyaux cérébelleux :
- le noyau fastigial est situé dans le vermis à proximité du toit du 4e ventricule ;
- les noyaux interposés, globuleux et emboliformes sont dans la partie la plus médiale de l'hémisphère ;
- les noyaux dentés sont situés plus latéralement enchâssés dans la substance blanche.
Anatomie de la moelle spinale
Fabrice Vuillier et Laurent Tatu
La moelle spinale constitue la partie inférieure du système nerveux central. Entièrement située dans le canal vertébral, elle donne naissance sur toute sa hauteur aux nerfs spinaux qui empruntent les foramens intervertébraux. Elle est le lieu de passage des voies ascendantes et descendantes et elle est le siège des arcs réflexes.
Comme toutes les structures appartenant au système nerveux central, la moelle spinale est constituée de substance grise, de substance blanche et de liquide cérébrospinal.
Configuration extérieure (figures 1.17 et 1.18)
La moelle spinale a la forme d'un tube cylindrique aplati d'avant en arrière. Elle possède une extrémité supérieure, qui fait suite au bulbe (moelle allongée) du tronc cérébral, et elle se termine par une extrémité inférieure arrondie appelée cône terminal rattachée au coccyx par le filum terminale.
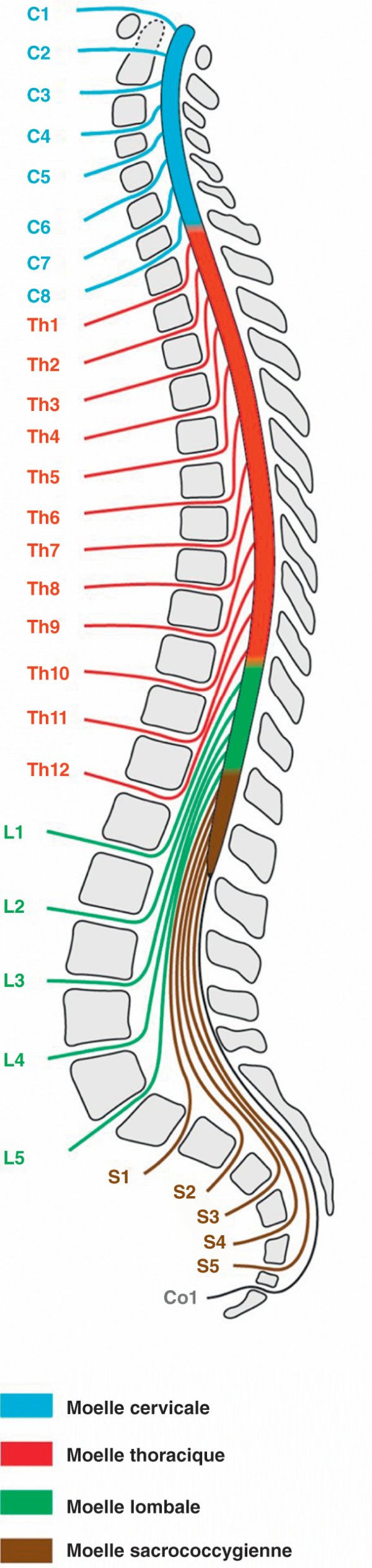
Figure 1.17
Situation de la moelle spinale.
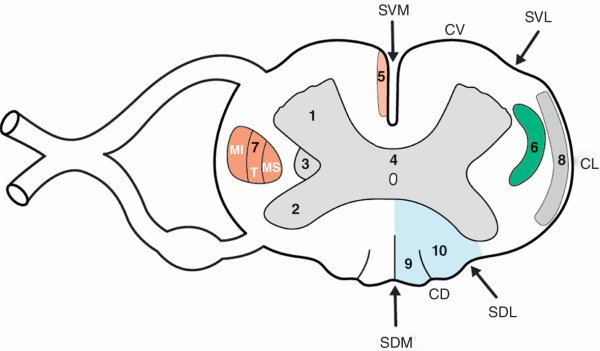
Figure 1.18
Coupe horizontale de la moelle spinale.
1. Corne ventrale de la moelle spinale. 2. Corne dorsale de la moelle spinale. 3. Corne latérale de la moelle spinale. 4. Substance grise intermédiaire et canal central de l'épendyme. 5. Faisceau corticospinal direct. 6. Faisceau spinothalamique. 7. Faisceau corticospinal croisé. 8. Faisceaux spinocérébelleux. 9. Faisceau gracile. 10. Faisceau cunéiforme. CD : cordon dorsal ; CL : cordon latéral ; CV : cordon ventral ; MI : membre inférieur ; MS : membre supérieur ; SDL : sillon dorsal latéral ; SDM : sillon dorsal médian ; SVL : sillon ventral latéral ; SVM : sillon ventral médian ; T : tronc.
La moelle spinale présente deux renflements correspondant à une plus grande densité de neurones destinés à l'innervation des membres : le renflement cervical est à l'origine des nerfs spinaux destinés aux membres supérieurs, et le renflement lombal à l'origine de ceux destinés aux membres inférieurs. Entre les renflements, se place la moelle thoracique dont le diamètre est plus réduit.
À sa surface, la moelle spinale est parcourue par des sillons verticaux dont le plus profond est le sillon ventral médian (fissure). Ces sillons délimitent des cordons pairs :
- cordon ventral, situé entre le sillon ventral médian et le sillon ventral latéral ;
- cordon latéral, situé entre les sillons ventral latéral et dorsal latéral ;
- cordon dorsal, situé entre le sillon dorsal latéral et le sillon dorsal médian.
Les sillons ventral latéral et dorsal latéral correspondent respectivement aux zones d'émergence des radicelles dont l'union forme les racines ventrales et dorsales des nerfs spinaux.
Selon la loi de la métamérisation, la moelle spinale est constituée d'un empilement de segments identiques (métamères) émettant chacun une paire de nerfs spinaux responsables d'un territoire cutané (dermatome), d'un territoire musculaire (myotome) et d'un territoire viscéral. La moelle spinale émet 31 paires de nerfs spinaux, chacun constitué par l'union d'une racine ventrale motrice et d'une racine dorsale sensitive.
Il est habituel de distinguer :
- la moelle cervicale, à l'origine de huit paires de nerfs spinaux cervicaux (C1 à C8) ;
- la moelle thoracique, à l'origine de douze paires de nerfs spinaux thoraciques (Th1 à Th12) ;
- la moelle lombale, à l'origine de cinq paires de nerfs spinaux lombaux (L1 à L5) ;
- la moelle sacrale, à l'origine de cinq paires de nerfs spinaux sacraux (S1 à S5) ;
- la moelle coccygienne, à l'origine d'une paire de nerfs spinaux coccygiens (Co1).
En moyenne la moelle spinale mesure 1 cm de diamètre (1,5 cm au niveau des renflements) et elle a une longueur correspondant à 25 % de la taille de l'individu.
Situation (cf. figure 1.17)
les différentes courbures. Elle n'occupe que les deux tiers de la longueur du canal vertébral en s'étendant habituellement du foramen magnum jusqu'à la hauteur du disque intervertébral séparant les 1re et 2e vertèbres lombales.
La projection vertébrale habituelle de la moelle spinale est la suivante :
- la moelle cervicale s'étend de la 1re à la 7e vertèbre cervicale ;
- la moelle thoracique s'étend de la 7e vertèbre cervicale à la 10e vertèbre thoracique ;
- la moelle lombale s'étend de la 10e à la 12e vertèbre thoracique ;
- les moelles sacrale et coccygienne s'étendent de la 12e vertèbre thoracique à la 2e vertèbre lombale.
Conséquence de cette différence morphologique entre le contenant (canal vertébral) et le contenu (moelle spinale), les trajets des racines et des nerfs spinaux deviennent de plus en plus obliques dans la partie inférieure du canal vertébral. En dessous du niveau du cône terminal, les racines lombales et sacrales disposées verticalement forment la queue de cheval à l'intérieur du canal vertébral.
Configuration intérieure (cf. figure 1.18)
Sur toute sa hauteur, la moelle spinale a une configuration intérieure uniforme. La substance grise est située au centre, autour du canal central de l'épendyme rempli de liquide cérébrospinal. La substance blanche est située en périphérie
Substance grise
Elle est organisée en une colonne centrale qui contient les corps cellulaires des neurones. Elle prend la forme d'un papillon aux ailes déployées avec de chaque côté :
- une corne ventrale, motrice, large, contenant les corps cellulaires des motoneurones alpha destinés à l'innervation des muscles striés ;
- une corne dorsale, sensitive, plus étroite, contenant les corps cellulaires des deutoneurones des voies de la sensibilité protopathique.
À l'étage de la moelle thoracique, une corne latérale appartenant au contingent sympathique du système nerveux autonome s'interpose entre les cornes ventrale et dorsale. Reliant les cornes ventrales et dorsales, la substance grise intermédiaire entoure le canal central de l'épendyme et comprend essentiellement des interneurones.
Substance blanche
Elle est constituée des axones myélinisés des neurones appartenant aux voies ascendantes, descendantes ainsi qu'aux voies d'association. Elle entoure la substance grise et s'organise en cordons :
- le cordon ventral est situé entre le sillon ventral médian et le sillon ventral latéral. Il contient notamment le faisceau corticospinal direct (motricité volontaire) qui ne représente qu'un faible contingent des fibres corticospinales ;
- le cordon latéral est situé entre le sillon ventral latéral et le sillon dorsal latéral. Il contient notamment les faisceaux spinothalamiques ventral et dorsal (sensibilité protopathique) et le faisceau corticospinal croisé (motricité volontaire). Ce dernier est organisé selon une somatotopie avec, de dedans en dehors, les fibres destinées aux membres supérieurs, puis au tronc et aux membres inférieurs. Il contient également les faisceaux spinocérébelleux ;
- le cordon dorsal est situé entre les sillons dorsal latéral et dorsal médian. Il contient les faisceaux véhiculant la sensibilité épicritique : faisceaux gracile en dedans (membres inférieurs) et cunéiforme en dehors (membres supérieurs).
Vascularisation artérielle (figure 1.19)
La moelle spinale est vascularisée par des artères qui perforent la substance nerveuse à partir d'un réseau anastomotique pie-mérien plaqué à la surface de la moelle. Ce réseau est alimenté par un système longitudinal, constitué de l'artère spinale ventrale et des artères spinales dorsales droite et gauche, et par un système transversal, constitué des artères segmentaires.
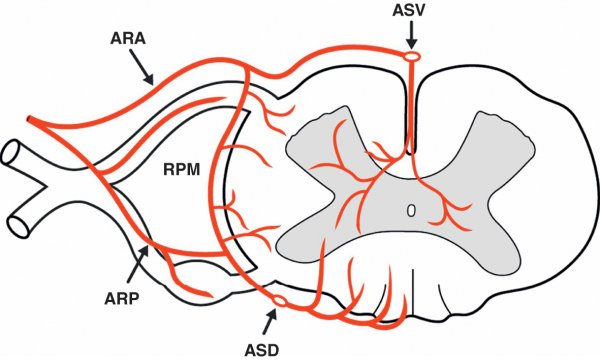
Figure 1.19
Organisation de la vascularisation artérielle de la moelle spinale.
ARA : artère radiculaire antérieure ; ARP : artère radiculaire postérieure ; ASD : artère spinale dorsale ; ASV : artère spinale ventrale ; RPM : réseau pie-mérien.
Système longitudinal
L'artère spinale ventrale, située dans le sillon ventral médian, naît de la convergence de deux rameaux nés chacun des artères vertébrales. Elle possède un calibre maximal au niveau du renflement cervical et plus réduit au niveau de la moelle thoracique. L'artère spinale ventrale est à l'origine de branches qui perforent la moelle spinale à travers le sillon ventral médian pour vasculariser la substance grise de chaque côté de la ligne médiane. Elle donne également des branches participant au riche réseau artériel pie-mérien situé à la surface des cordons.
Les artères spinales dorsales, plus grêles, naissent le plus souvent des artères vertébrales et se placent chacune le long du sillon dorsal latéral de la moelle jusqu'au cône terminal où elles forment avec l'artère spinale ventrale une boucle anastomotique. Par de nombreuses branches, elles alimentent le réseau anastomotique pie-mérien.
Système transversal
Ce système est constitué des artères radiculaires antérieures et postérieures qui, portées par les racines des nerfs spinaux, viennent s'aboucher sur le système longitudinal. Au niveau des renflements, ces artères sont de plus gros volume et participent à la vascularisation de la moelle spinale en abordant directement le cordon latéral. C'est le cas notamment de l'artère du renflement lombal (artère d'Adamkiewicz).
Système liquidien
Fabrice Vuillier et Laurent Tatu
Le liquide cérébrospinal est un liquide propre au système nerveux central dont la composition est proche de celle du plasma. Sécrété par les plexus choroïdes des ventricules encéphaliques, il est résorbé par les granulations arachnoïdiennes. C'est un liquide transparent « eau de roche » qui circule à l'intérieur du système ventriculaire encéphalique (système liquidien interne) et dans les espaces subarachnoïdiens délimités par les méninges (système liquidien externe). Il assure le transport d'éléments nutritifs pour le tissu nerveux et l'élimination des produits du métabolisme et il possède un rôle mécanique de protection du système nerveux central.
Méninges
Ce sont des tissus qui enveloppent et protègent de façon concentrique le système nerveux central. De la superficie à la profondeur se placent successivement :
- la dure-mère, méninge de protection, épaisse et résistante (pachyméninge) ;
- l'arachnoïde, méninge de glissement, plus mince (leptoméninge), qui double en profondeur la dure-mère ;
- la pie-mère, également mince (leptoméninge), méninge nourricière véritable porte-vaisseaux, intimement appliquée à la surface du tissu nerveux.
Autour et entre eux, les méninges délimitent les espaces extradural (épidural), subdural et subarachnoïdien.
On distingue les méninges spinales, qui entourent la moelle spinale, et les méninges encéphaliques, qui entourent l'encéphale.
Disposition générale des méninges spinales (figure 1.20)
La dure-mère spinale enveloppe la moelle spinale en formant un sac allongé dans le canal vertébral, du foramen magnum jusqu'à hauteur de la 3e vertèbre sacrale. Les racines des nerfs spinaux traversent la dure-mère qui les accompagne en formant un manchon jusqu'au foramen intervertébral. Elle est séparée des parois du canal vertébral par l'espace extradural (épidural) contenant principalement de la graisse et des plexus veineux. C'est dans cet espace que s'effectue l'injection lors d'une anesthésie épidurale.
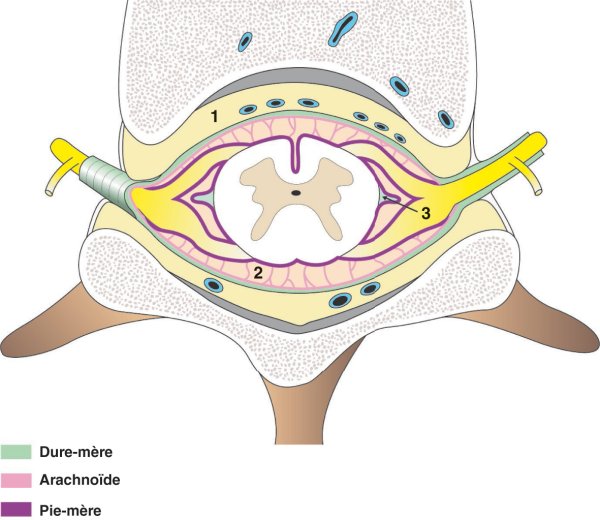
Figure 1.20
Disposition des méninges spinales.
1. Espace extradural (épidural). 2. Espace subarachnoïdien. 3. Ligament dentelé.
L'arachnoïde spinale recouvre toute la face profonde de la dure-mère. Elle est séparée de la pie-mère par l'espace subarachnoïdien dans lequel se situe le liquide cérébrospinal que l'on prélève lors de la ponction lombaire. Cet espace est cloisonné par la présence de trabéculations qui s'attachent à la pie-mère.
La pie-mère est appliquée contre la surface de la moelle spinale et s'insinue dans les sillons. Elle émet deux prolongements latéraux formant les ligaments dentelés qui contribuent au maintien de la moelle spinale.
Disposition générale des méninges encéphaliques (figures 1.21 et 1.22)
La dure-mère encéphalique enveloppe l'encéphale en adhérant aux os du crâne à l'exception de l'os temporal au niveau d'une zone décollable (de Gérard Marchant), siège privilégié de l'hématome extradural. Elle envoie des expansions entre les deux hémisphères cérébraux (faux du cerveau) et entre le cerveau en haut et le cervelet en bas (tente du cervelet).
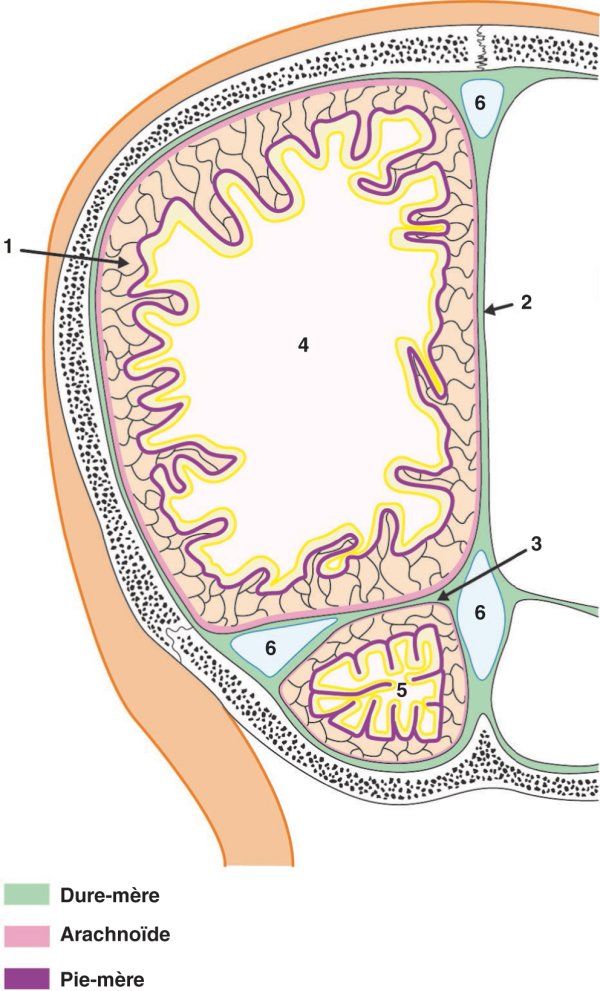
Figure 1.21
Disposition des méninges encéphaliques.
1. Espace subarachnoïdien. 2. Faux du cerveau. 3. Tente du cervelet. 4. Cerveau. 5. Cervelet. 6. Sinus duraux.
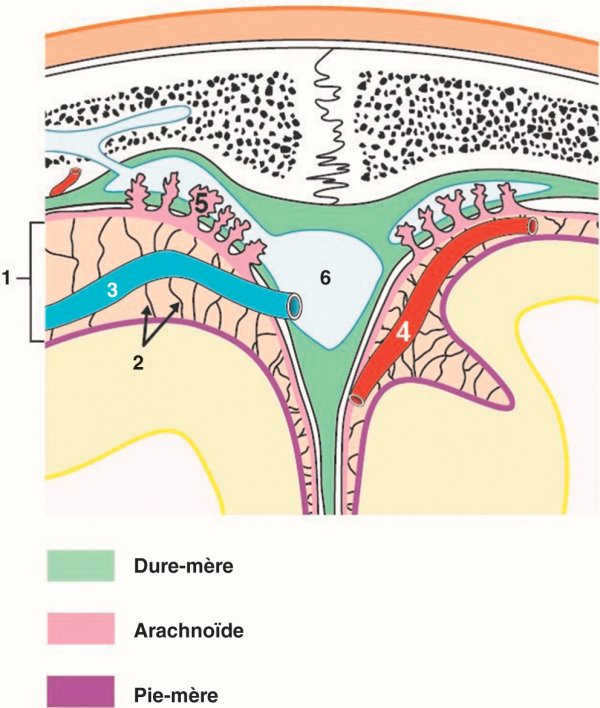
Figure 1.22
Espace subarachnoïdien encéphalique.
1. Espace subarachnoïdien. 2. Trabéculations arachnoïdiennes. 3. Veine corticale. 4. Artère corticale. 5. Granulations arachnoïdiennes. 6. Sinus dural.
En se dédoublant, la dure-mère encéphalique renferme les sinus duraux qui drainent l'ensemble du sang veineux de l'encéphale vers les veines jugulaires internes.
L'arachnoïde se place en dedans de la dure-mère dont elle est séparée par un espace virtuel contenant des veines (espace subdural). Entre l'arachnoïde et la pie-mère, se place l'espace subarachnoïdien dans lequel circule le liquide cérébrospinal et où sont placées les trabéculations arachnoïdiennes qui rejoignent la pie-mère. L'arachnoïde émet des villosités situées dans les sinus veineux et qui sont chargées de résorber le liquide cérébrospinal vers la circulation veineuse. La pie-mère est appliquée contre le cortex du cerveau dont elle suit les sillons.
Ventricules encéphaliques (figure 1.23)
Ils constituent le système liquidien interne et forment un ensemble de cavités situées en profondeur de l'encéphale. Remplis de liquide cérébrospinal, ils contiennent également les plexus choroïdes.
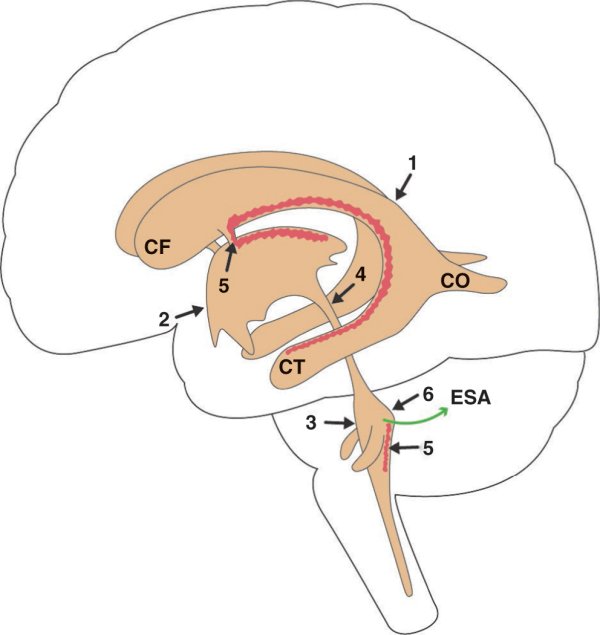
Figure 1.23
Ventricules encéphaliques.
1. Ventricule latéral gauche. 2. 3e ventricule. 3. 4e ventricule. 4. Aqueduc du mésencéphale. 5. Plexus choroïdes. 6. Orifice du 4e ventricule. CF : corne frontale du ventricule latéral ; CO : corne occipitale du ventricule latéral ; CT : corne temporale du ventricule latéral ; ESA : espace subarachnoïdien.
Les ventricules encéphaliques sont représentés par les ventricules latéraux, le 3e ventricule (ventricule diencéphalique) et le 4e ventricule (ventricule rhombencéphalique). Ces ventricules communiquent entre eux ainsi qu'avec les espaces subarachnoïdiens et le canal central de l'épendyme.
Les ventricules latéraux sont pairs, symétriques et situés dans la partie profonde de chaque hémisphère. Arciformes à concavité antéro-inférieure, ils possèdent un corps et trois expansions sous forme de cornes frontale, temporale et occipitale. Les ventricules latéraux communiquent avec le 3e ventricule dans lequel il déverse le liquide cérébrospinal à travers les foramens interventriculaires (de Monro).
Le 3e ventricule est une cavité impaire et médiane située dans le diencéphale. Il communique avec le 4e ventricule par l'aqueduc du mésencéphale. Un simple rétrécissement de ce canal déjà étroit est à l'origine d'un certain nombre d'hydrocéphalies.
De forme triangulaire, le 4e ventricule est une cavité impaire et médiane creusant le rhombencéphale. Sa paroi antérieure, correspondant au plancher du 4e ventricule (fosse rhomboïde), est constituée de la face dorsale du bulbe et du pont et son toit est constitué du vermis cérébelleux et des voiles médullaires supérieur et inférieur. À sa partie inférieure, le toit est percé d'un orifice permettant le passage du liquide cérébrospinal du système ventriculaire vers les espaces subarachnoïdiens.
Vascularisation artérielle de l'encéphale
Fabrice Vuillier et Laurent Tatu
L'encéphale ne représente que 2 % du poids du corps mais il absorbe près de 20 % de la consommation énergétique globale. Sa vascularisation artérielle est donc essentielle afin de lui assurer un apport métabolique adapté.
Le système artériel de l'encéphale est organisé selon trois niveaux (figure 1.24) :
- les voies artérielles d'apport, constituées par trois piliers : les artères carotides internes en avant et l'axe vertébrobasilaire en arrière ;
- le cercle artériel de la base du cerveau (polygone de Willis), système d'anastomoses entre les voies d'apport et base de lancement des artères cérébrales ;
- les artères cérébrales chargées de vasculariser les parties superficielles et profondes des hémisphères.
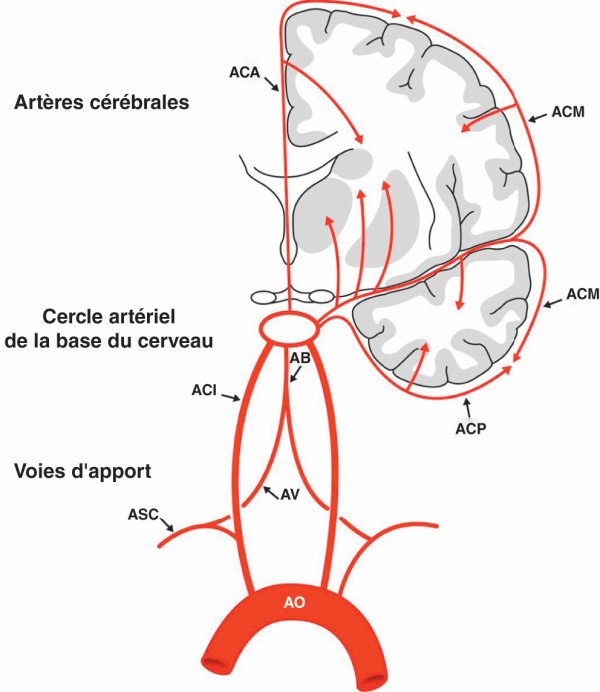
Figure 1.24
Les trois niveaux du système artériel encéphalique.
AB : artère basilaire ; ACA : artère cérébrale antérieure ; ACI : artère carotide interne ; ACM : artère cérébrale moyenne ; ACP : artère cérébrale postérieure ; AO : aorte ; ASC : artère subclavière ; AV : artère vertébrale.
Voies artérielles d'apport (figure 1.25)
Artère carotide interne
À droite comme à gauche, l'artère carotide interne est la branche de division postérolatérale de l'artère carotide commune. Elle naît à hauteur de la 4e vertèbre cervicale et son trajet s'étend jusqu'au-dessus du toit du sinus caverneux.
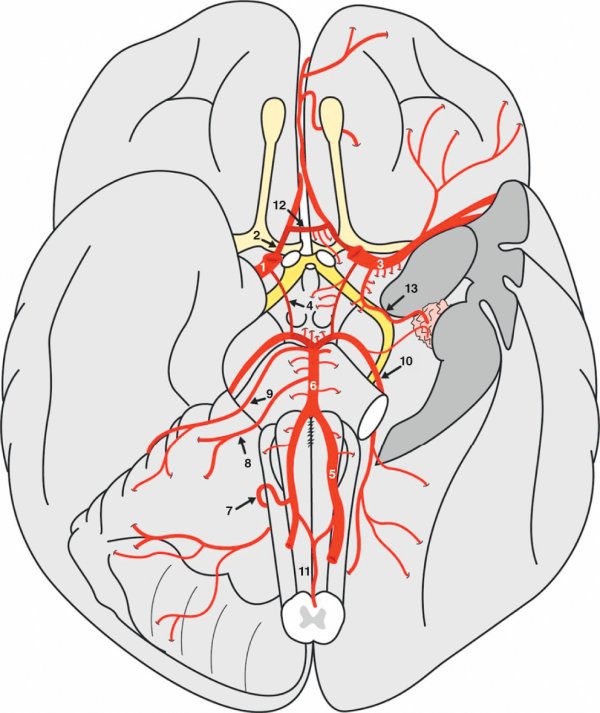
Figure 1.25
Les artères de l'encéphale (vue inférieure de l'encéphale).
1. Artère carotide interne. 2. Artère cérébrale antérieure. 3. Artère cérébrale moyenne. 4. Artère communicante postérieure. 5. Artère vertébrale. 6. Artère basilaire. 7. Artère cérébelleuse inférieure. 8. Artère cérébelleuse moyenne. 9. Artère cérébelleuse supérieure. 10. Artère cérébrale postérieure. 11. Artère spinale antérieure. 12. Artère communicante antérieure. 13. Artère choroïdienne antérieure.
Au cours de son trajet, elle a d'abord une portion cervicale, au cours de laquelle son trajet ascendant est rectiligne, l'amenant au niveau de la base du crâne, à proximité du processus styloïde. Puis elle traverse l'os temporal dans le canal carotidien (portion pétreuse) selon un trajet en baïonnette.
Dans la boîte crânienne, l'artère carotide interne traverse le sinus caverneux de bas en haut (portion caverneuse) selon un trajet en forme de S italique (siphon carotidien), puis elle perfore son toit. À la sortie du sinus caverneux, elle se termine en se divisant en ses quatre branches terminales : les artères cérébrales antérieure, cérébrale moyenne, choroïdienne antérieure et communicante postérieure.
Axe vertébrobasilaire
Les artères vertébrales droite et gauche sont des branches collatérales des artères subclavières droite et gauche. Elles cheminent dans le canal transversaire des six premières vertèbres cervicales puis entrent dans la boîte crânienne par le foramen magnum. Elles se placent sur la face ventrale du bulbe (moelle allongée) et fusionnent à hauteur du sillon bulbopontique pour former l'artère basilaire. Avant de fusionner, chaque artère vertébrale donne une artère cérébelleuse inférieure et une artère spinale antérieure.
En se plaçant sur la face ventrale du pont, l'artère basilaire monte jusqu'au sillon pontomésencéphalique où elle se termine en ses deux branches terminales, les artères cérébrales postérieures droite et gauche. L'artère basilaire donne comme principales branches collatérales les artères cérébelleuses moyenne et supérieure.
Cercle artériel de la base du cerveau (figures 1.25 et 1.26)
Situé à la face inférieure du cerveau, le cercle artériel de la base du cerveau a la forme d'un polygone dont les côtés sont constitués par :
- en avant, les deux segments proximaux (A1) des artères cérébrales antérieures unis par l'artère communicante antérieure ;
- en arrière, les deux segments proximaux (P1) des artères cérébrales postérieures ;
- latéralement, les deux artères communicantes postérieures.
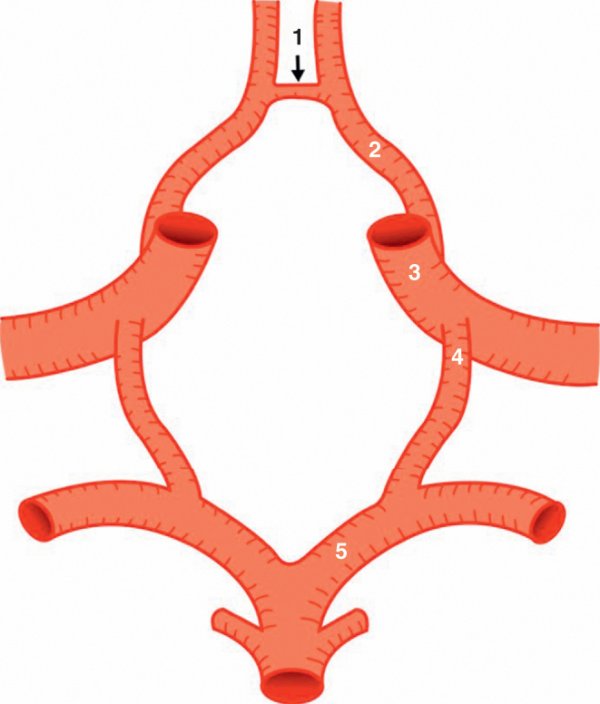
Figure 1.26
Le cercle artériel de la base du cerveau.
1. Artère communicante antérieure. 2. Segment proximal (A1) de l'artère cérébrale antérieure. 3. Artère carotide interne. 4. Artère communicante postérieure. 5. Segment proximal (P1) de l'artère cérébrale postérieure.
Le cercle artériel de la base du cerveau est le siège de nombreuses variations morphologiques (absence d'une artère constitutive ou de calibres artériels variables). L'artère communicante postérieure est l'artère soumise aux plus nombreuses variations.
Le cercle artériel de la base du cerveau est la véritable base de lancement des artères cérébrales antérieure, moyenne et postérieure. Les artères constitutives du cercle artériel de la base du cerveau sont aussi à l'origine de branches collatérales (artères perforantes profondes) qui, en passant à travers les substances perforées antérieures, latérales et postérieures, participent à la vascularisation des structures situées en profondeur des hémisphères.
Artères cérébrales (figures 1.27 et 1.28)
La circulation artérielle cérébrale est organisée en deux systèmes alimentés essentiellement par les artères cérébrales antérieure, moyenne, postérieure et choroïdienne antérieure.
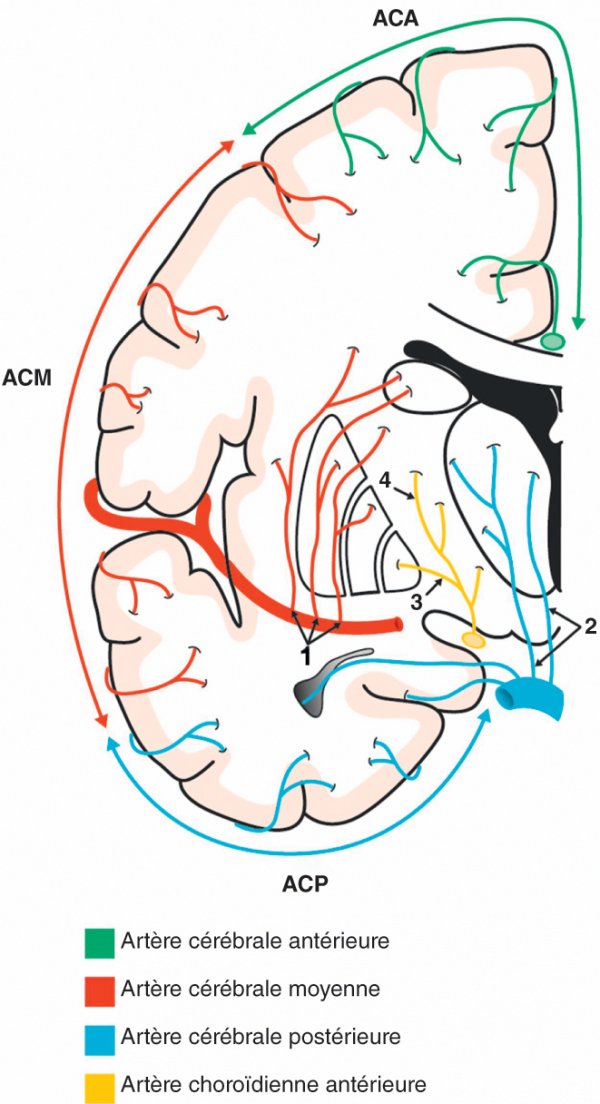
Figure 1.27
Artères corticales et perforantes profondes du cerveau.
1. Artères lenticulostriées. 2. Artères thalamiques. 3. Artères du globus pallidus médial. 4. Artères du bras postérieur de la capsule interne.
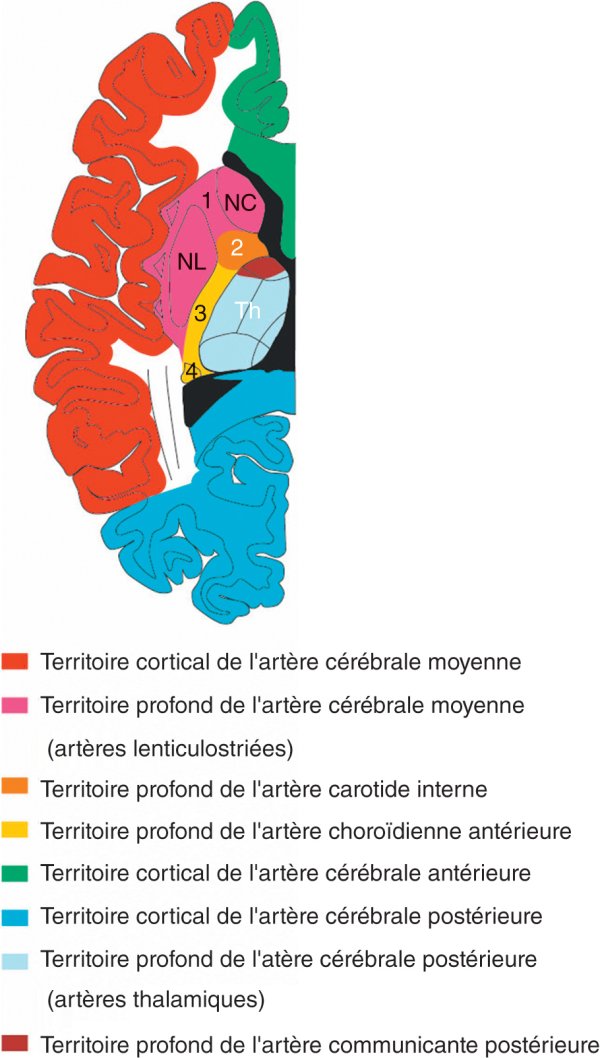
Figure 1.28
Coupe schématique des principaux territoires artériels du cerveau.
1. Bras antérieur de la capsule interne. 2. Genou de la capsule interne. 3. Bras postérieur de la capsule interne. 4. Queue du noyau caudé. NC : noyau caudé ; NL : noyau lentiforme ; Th : thalamus.
Le système superficiel assure la vascularisation des territoires corticaux des hémisphères. Il est constitué d'artères corticales, leptoméningées ou piales, qui forment un réseau artériel anastomotique à la surface du cerveau. Ces artères envoient des branches qui pénètrent à angle droit le cortex et atteignent la substance blanche sous-jacente.
Le système profond assure la vascularisation des territoires profonds des hémisphères, c'est-à-dire le diencéphale et les noyaux de la base. Il est alimenté par les artères perforantes profondes issues du cercle artériel de la base du cerveau ou de la partie proximale de ses branches.
Artère cérébrale antérieure
Le territoire de vascularisation des artères corticales de l'artère cérébrale antérieure comprend les deux tiers médiaux de la face inférieure du lobe frontal et les faces médiales des lobes frontal et pariétal. Elles vascularisent aussi les parties supéromédiales des gyrus frontal supérieur, précentral et post-central.
Le territoire de vascularisation des artères perforantes profondes de l'artère cérébrale antérieure comprend essentiellement la partie antéro-inférieure de la tête du noyau caudé, la partie antérieure et inférieure du bras antérieur de la capsule interne, et la partie antérieure du globus pallidus.
Artère cérébrale moyenne
Le territoire de vascularisation des artères corticales issues de l'artère cérébrale moyenne comprend la partie latérale de la face inférieure du lobe frontal, la totalité de la face latérale des lobes frontal et pariétal ainsi que l'insula, les opercules et la face latérale du lobe temporal.
Le territoire de vascularisation des artères perforantes profondes de l'artère cérébrale moyenne (artères lenticulostriées) comprend la partie supérieure de la tête et le corps du noyau caudé, le putamen, la partie latérale du globus pallidus, la partie supérieure du bras antérieur, du genou et du bras postérieur de la capsule interne.
Artère cérébrale postérieure
Le territoire de vascularisation des artères corticales de l'artère cérébrale postérieure est constitué de la partie inféromédiale du lobe temporal située entre le sillon temporal inférieur et la fissure pariéto-occipitale, à l'exception du pôle temporal.
Le territoire de vascularisation des artères perforantes profondes de l'artère cérébrale postérieure comprend la majeure partie du tronc cérébral, du thalamus et des corps géniculés.
Artère choroïdienne antérieure
Le territoire de vascularisation des artères corticales de l'artère choroïdienne
Le territoire de vascularisation des artères perforantes profondes de l'artère choroïdienne antérieure comprend la partie ventrale des deux tiers postérieurs du bras postérieur de la capsule interne, une partie du tractus optique, la partie latérale du corps géniculé latéral, la partie inférieure de la queue du noyau caudé et la partie médiale du globus pallidus.
Vascularisation artérielle du tronc cérébral et du cervelet (figure 1.29)
La vascularisation artérielle du tronc cérébral et du cervelet est assurée par des branches provenant de l'axe vertébrobasilaire. Ces artères ont une longueur variable mais elles pénètrent le tronc cérébral à des points fixes et alimentent les territoires antéromédial, antérolatéral, latéral et postérieur du tronc cérébral. Ces territoires, qui sont constants aux trois étages du tronc cérébral, sont alimentés par des pédicules artériels qui varient en fonction des différents niveaux du tronc cérébral.
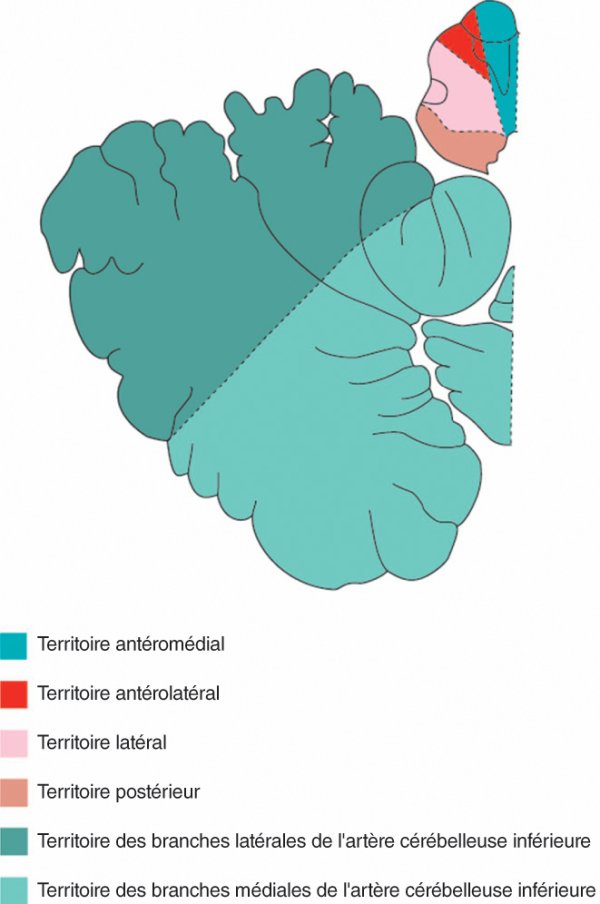
Figure 1.29
Coupe schématique des territoires artériels du bulbe et du cervelet.
Le cervelet est vascularisé par les artères cérébelleuses inférieures, moyennes et supérieures qui se divisent chacune en une branche médiale et latérale. Au cours de leur trajet, les artères cérébelleuses participent, par leurs branches médiales, à la vascularisation du tronc cérébral.
Anatomie fonctionnelle des voies de la sensibilité
Fabrice Vuillier et Laurent Tatu
La sensibilité somatique (somesthésie) et les sensibilités spéciales (organes des sens) permettent aux organismes vivants de réagir aux stimulations externes et internes. Concernant la sensibilité somatique, la classification la plus pertinente, sur le plan anatomique et fonctionnel pour une bonne utilisation en pratique clinique, distingue les sensibilités épicritique, protopathique et la sensibilité proprioceptive inconsciente :
- la sensibilité épicritique regroupe la perception du tact fin et conscient qui permet la discrimination et la localisation des informations et la proprioception consciente (pallesthésie, kinesthésie). Elle emprunte la voie anatomique, dite lemniscale, qui passe par le lemniscus médial du tronc cérébral. Elle est composée de fibres nerveuses myélinisées et de gros calibre permettant une conduction rapide et une bonne discrimination spatiale ;
- la sensibilité protopathique regroupe la perception du tact grossier, de la température et la douleur. Elle emprunte la voie anatomique dite extralemniscale et voyage dans le faisceau spinothalamique. Elle est composée de fibres nerveuses peu ou non myélinisées, de petit calibre, à l'origine d'une vitesse de conduction plus lente et d'une moins bonne discrimination spatiale ;
- la sensibilité proprioceptive inconsciente est véhiculée par une autre voie anatomique constituée des faisceaux spinocérébelleux. Elle est chargée d'informer le cervelet pour sa fonction de contrôle du mouvement.
Les voies protopathique et épicritique ont des caractéristiques générales communes :
- elles sont ascendantes et croisées ;
- elles transportent l'information du récepteur, situé en périphérie, jusqu'au cortex cérébral ;
- elles sont organisées selon un réseau de trois neurones (protoneurone, deutoneurone, neurone thalamocortical) dans lequel le relais entre le 2e et le 3e neurone s'effectue au niveau du thalamus.
Sensibilité épicritique (figure 1.30)
Le premier neurone de cette voie possède un corps cellulaire situé dans le ganglion spinal de la racine dorsale du nerf spinal. Ses dendrites, au contact de récepteurs musculotendineux et d'autres situés dans le derme, sont transportées par les nerfs du système nerveux périphérique, puis par la racine dorsale des nerfs spinaux. L'axone pénètre dans la moelle spinale et alimente les faisceaux gracile (axones sacrés et lombaires) et cunéiforme (axones thoraciques et cervicaux) du cordon dorsal de la moelle spinale en déterminant une somatotopie. L'axone du 1er neurone établit un relais avec le 2e neurone dont le corps cellulaire se situe au niveau des noyaux gracile et cunéiforme situés dans la partie dorsale du bulbe fermé.
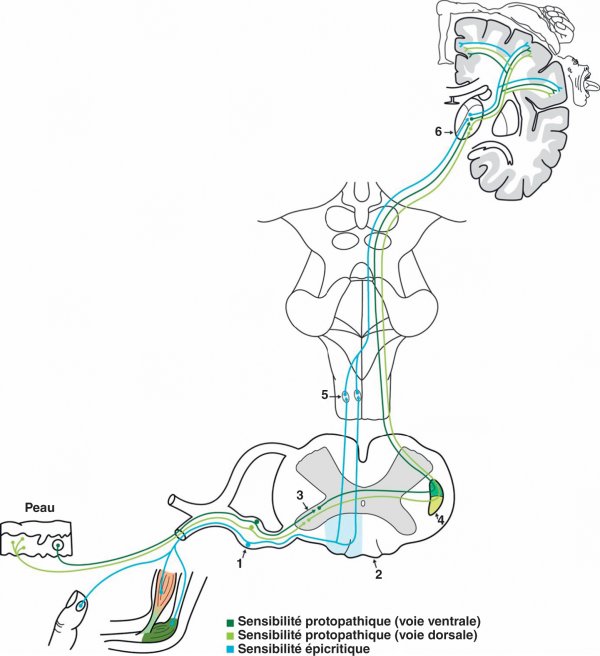
Figure 1.30
Voies sensitives.
1. Ganglion spinal. 2. Cordons dorsaux de la moelle spinale. 3. Corne dorsale de la moelle spinale. 4. Faisceau spinothalamique. 5. Lemniscus médial. 6. Thalamus ventral.
L'axone du 2e neurone croise la ligne médiane en formant le lemniscus médial et parcourt les trois étages du tronc cérébral pour gagner la partie ventrale (pars oralis) du noyau ventro-postéro-latéral du thalamus où s'établit le relais avec le neurone thalamocortical. Ce noyau est organisé selon une somatotopie avec de dedans en dehors les fibres du membre supérieur, du tronc et du membre inférieur (figure 1.31). Les informations provenant de la face sont transportées par le faisceau trigéminothalamique qui se projette sur le noyau ventro-postéro-médial du thalamus.
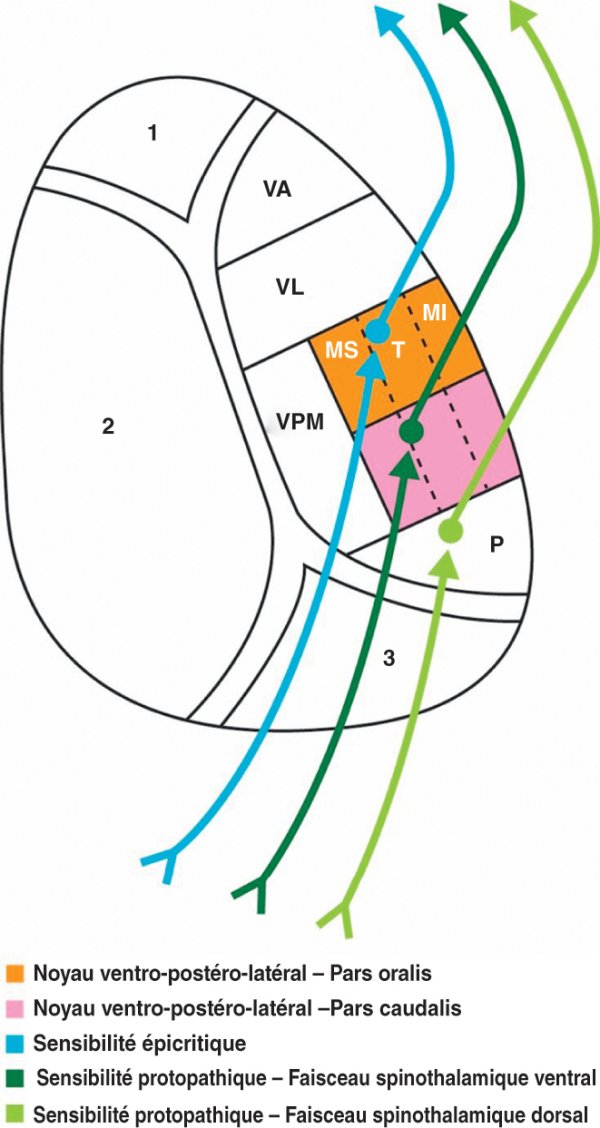
Figure 1.31
Relais thalamiques des voies sensitives.
1. Groupe nucléaire antérieur. 2. Groupe nucléaire médial. 3. Groupe nucléaire postérieur. MI : membre inférieur ; MS : membre supérieur ; P : noyau postérieur ; T : tronc ; VA : noyau ventral antérieur ; VL : noyau ventral latéral ; VPM : noyau ventral postéromédian.
L'axone du neurone thalamocortical emprunte le bras postérieur de la capsule interne pour se projeter sur le gyrus post-central du lobe pariétal (aire somatosensible primaire) organisé selon une somatotopie décrivant l'homonculus sensitif, superposable à l'homonculus moteur (de Penfield).
Sensibilité protopathique (cf. figure 1.30)
Le 1er neurone de cette voie possède un corps cellulaire situé dans le ganglion spinal de la racine dorsale du nerf spinal. Les dendrites, au contact de récepteurs sensibles au tact grossier, à la température et à la douleur, sont transportées par les nerfs du système nerveux périphérique puis par la racine dorsale des nerfs spinaux. L'axone gagne la substance grise de la corne dorsale au sein de laquelle s'effectue le relais avec le 2e neurone. L'axone du 2e neurone franchit la ligne médiane et participe à la constitution des faisceaux spinothalamique ventral (tact grossier, température, douleur) et spinothalamique dorsal (localisation précise de la douleur) qui se placent dans le cordon latéral de la moelle spinale. Le faisceau spinothalamique ventral gagne la partie dorsale (pars caudalis) du noyau ventro-postéro-latéral du thalamus et le faisceau spinothalamique dorsal se projette sur le noyau postérieur du thalamus où s'établissent les relais avec le 3e neurone (cf. figure 1.31). Le noyau ventro-postéro-latéral du thalamus est le siège d'une somatotopie où les fibres transportant des informations provenant du membre supérieur, du tronc puis du membre inférieur se disposent respectivement de dedans en dehors.
L'axone du neurone thalamocortical emprunte le bras postérieur de la capsule interne pour se projeter sur le gyrus post-central du lobe pariétal (aire somatosensible primaire) organisé selon une somatotopie décrivant l'homonculus sensitif.
Il existe également une voie spino-réticulo-thalamique satellite des voies spinothalamiques. C'est une voie multisynaptique dont les relais s'effectuent dans la substance réticulée du tronc cérébral ainsi que dans les noyaux intralaminaires du thalamus avant de se terminer au niveau du lobe limbique. Cette voie plus complexe intervient notamment dans la composante émotionnelle de la douleur.
De manière physiologique, le premier relais sur les voies protopathiques peut être soumis à un blocage du passage de l'influx douloureux par l'action inhibitrice d'une collatérale de l'axone du 1er neurone de la voie épicritique. Cette influence inhibitrice de la voie épicritique sur la voie protopathique permet un contrôle de la douleur et ce phénomène qui correspond au gate control est à la base de certaines thérapeutiques de neurostimulation (figure 1.32).
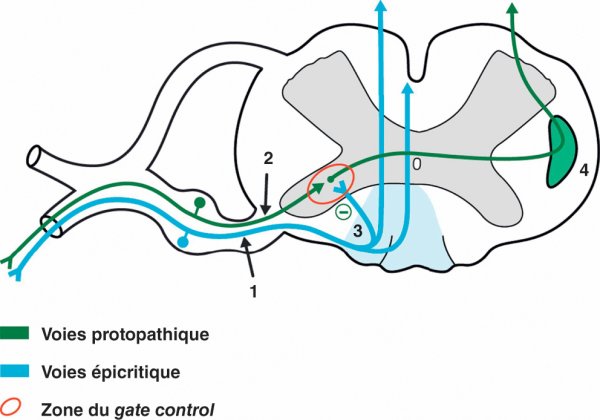
Figure 1.32
Voies sensitives dans la moelle spinale.
1. Premier neurone de la voie épicritique. 2. Premier neurone de la voie protopathique. 3. Collatérale du gate control. 4. Faisceau spinothalamique.
Anatomie fonctionnelle de la motricité
Laurent Tatu et Fabrice Vuillier
Voies motrices volontaires (figure 1.33)
Les voies de la motricité volontaire regroupent le faisceau corticospinal (anciennement dénommé « faisceau pyramidal ») et le faisceau corticonucléaire (appelé précédemment « faisceau géniculé »). Ces deux faisceaux ne constituent que la partie exécutive d'une organisation plus complexe de la motricité volontaire qui inclut également les ganglions de la, base et le cortex prémoteur pour la planification, le cervelet intermédiaire pour le contrôle du mouvement en cours, et le cervelet latéral pour la programmation du mouvement.
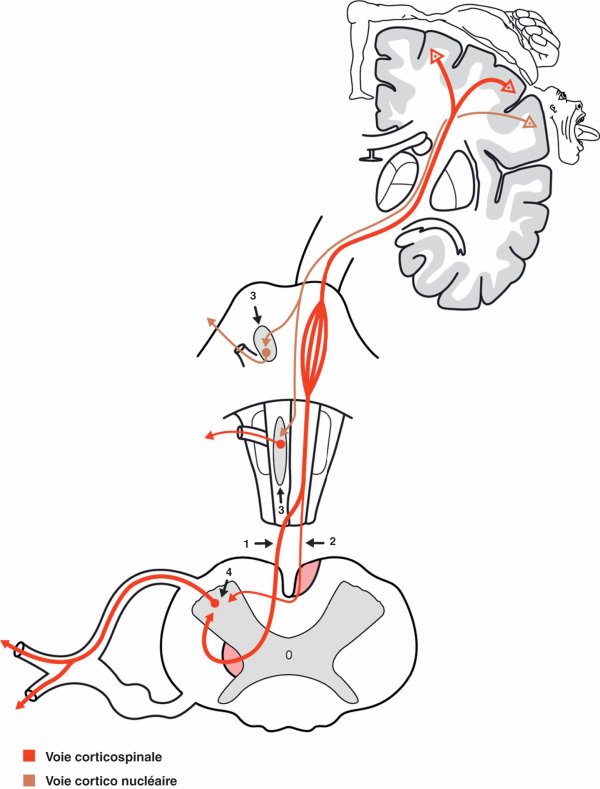
Figure 1.33
Voies motrices volontaires.
1. Faisceau corticospinal croisé. 2. Faisceau corticospinal direct. 3. Noyaux moteurs des nerfs crâniens. 4. Corne ventrale de la moelle spinale.
Les voies de la motricité volontaire correspondent à des voies simples, croisées, formées de deux neurones qui relient les centres corticaux et le système musculaire. Les faisceaux de la motricité volontaire traversent le système nerveux central : le centre ovale, la capsule interne, le tronc cérébral et, pour le faisceau corticospinal, la moelle spinale. Durant tout leur trajet, ils conservent une organisation somatotopique.
Aires corticales de la motricité
Les corps cellulaires des premiers neurones des voies corticospinale et corticonucléaire se placent de manière prépondérante dans le gyrus précentral qui constitue l'aire motrice primaire placée verticalement entre la fissure centrale en arrière et le sillon précentral. D'autres neurones moins nombreux se trouvent dans les aires corticales frontales prémotrices, sur la face latérale des circonvolutions F1, F2 et F3 qui interviennent dans les mouvements spécifiques des globes oculaires par exemple. D'autres aires prémotrices sont également présentes sur la face médiale de l'hémisphère en avant du lobule paracentral en formant l'aire motrice supplémentaire (AMS) impliquée dans la planification du mouvement et dans la coordination des mouvements complexes des membres. Un autre contingent plus réduit de neurones est présent dans le gyrus post-central, correspondant à l'aire sensitive primaire entre la fissure centrale et le sillon post-central.
Trajet
Les axones des premiers neurones traversent le centre ovale de l'hémisphère cérébral et constituent progressivement les faisceaux corticospinal et corticonucléaire qui traversent ensuite le bras postérieur de la capsule interne et pénètrent le pédoncule cérébral du mésencéphale.
Au niveau du pont, le faisceau corticospinal est dispersé en de multiples fascicules par la présence des noyaux du pont. Il se reconstitue en un tractus compact à hauteur du bulbe (medulla oblongata). À hauteur de la partie inférieure du bulbe, environ 80 % des axones de la voie corticospinale décussent pour former le tractus corticospinal latéral (faisceau corticospinal croisé) dans le cordon latéral de la moelle spinale. Les autres axones ne croisent pas la ligne médiane au niveau du bulbe et forment le tractus corticospinal ventral (faisceau corticospinal direct) dans le cordon ventral de la moelle spinale.
Les axones du 1er neurone du tractus corticospinal latéral font relais avec le deuxième neurone moteur dans la corne ventrale de la moelle spinale. Ceux du tractus corticospinal ventral décussent à chaque niveau segmentaire par la commissure antérieure de la moelle spinale pour également faire relais avec le 2e neurone moteur dans la corne ventrale. L'axone du 2e neurone moteur emprunte la racine ventrale du nerf spinal, puis le nerf spinal et ses branches ventrale et dorsale pour innerver les muscles des membres et du tronc.
Les axones constituant le faisceau corticonucléaire décussent à différents niveaux du tronc cérébral pour rejoindre les noyaux moteurs controlatéraux des nerfs crâniens. Ils font relais avec le 2e neurone moteur dont les axones empruntent les nerfs crâniens pour innerver les muscles à commande volontaire de l'extrémité cervicocéphalique. Le faisceau corticonucléaire s'amenuise progressivement dans la traversée du tronc cérébral pour finalement disparaître au niveau bulbaire. Certains noyaux moteurs des nerfs crâniens, comme ceux du nerf facial, reçoivent une afférence bilatérale des faisceaux corticonucléaires.
Somatotopie
Le cortex moteur du gyrus précentral est le siège d'une somatotopie. Chaque région de l'hémicorps controlatéral est représentée sur ce gyrus. Les régions du corps avec une grande densité d'innervation musculaire, comme la main ou la langue, ont une plus grande représentation corticale que les régions avec une plus faible densité d'innervation, comme le tronc. Cette représentation disproportionnée est appelée « homonculus moteur de Penfield ».
La représentation somatotopique se poursuit tout au long des voies motrices volontaires. Dans la traversée de la capsule interne, le faisceau corticonucléaire occupe le genou délimité, en dedans, par le corps du noyau caudé et le thalamus et, en dehors, par l'extrémité du globus pallidus. Il véhicule la partie de la voie motrice volontaire destinée à l'extrémité cervicocéphalique. Le faisceau corticospinal se place dans le bras postérieur de la capsule interne entre le thalamus en dedans et le globus pallidus et le putamen en dehors. Dans la partie motrice du bras postérieur de la capsule interne, la somatotopie du faisceau corticospinal s'organise de l'avant vers l'arrière : membre supérieur, tronc, membre inférieur (figure 1.34).
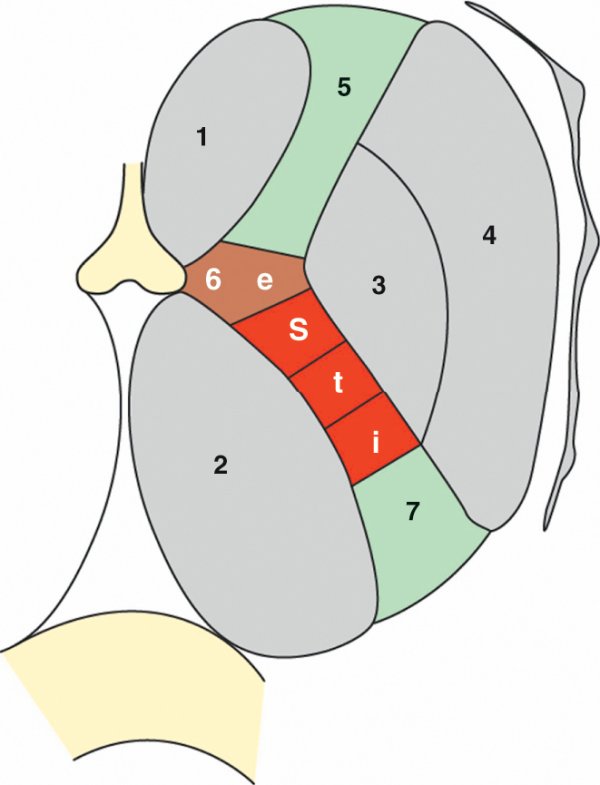
Figure 1.34
Somatotopie des voies motrices dans la capsule interne.
1. Noyau caudé. 2. Thalamus. 3. Globus pallidus. 4. Putamen. 5. Bras antérieur de la capsule interne. 6. Genou de la capsule interne. 7. Bras postérieur de la capsule interne. e : extrémité céphalique ; i : membre inférieur ; s : membre supérieur ; t : tronc.
Dans la moelle spinale, le faisceau corticospinal croisé est organisé de manière somatotopique. Les fibres destinées aux différents segments du corps se placent, de dedans en dehors : segments cervical, thoracique, lombal et sacral. Il existe également une organisation somatotopique dans la corne ventrale de la moelle spinale. Les neurones moteurs sont organisés en groupes cellulaires. Par exemple, au niveau segmentaire correspondant à l'innervation des membres, les groupes médiaux innervent les muscles de la partie proximale du membre et les groupes latéraux les muscles de la partie distale (figure 1.35).
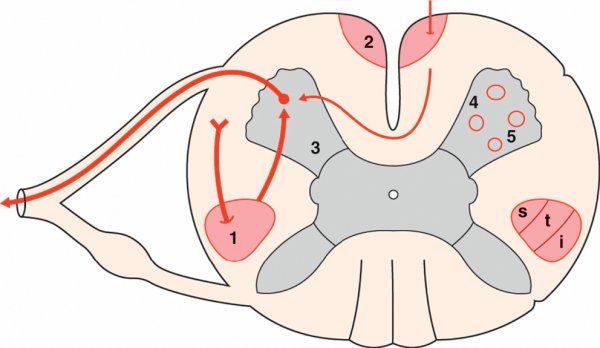
Figure 1.35
Voies motrices dans la moelle spinale.
1. Faisceau corticospinal croisé. 2. Faisceau corticospinal direct. 3. Corne ventrale de la moelle spinale. 4. Colonnes médiales de la corne ventrale. 5. Colonnes latérales de la corne ventrale. i : membre inférieur ; s : membre supérieur ; t : tronc.
Contrôle cérébelleux
La morphologie complexe du cervelet ne correspond pas à son organisation fonctionnelle. La division phylogénétique classique en archéo, paléo et néocérébellum n'a que peu d'intérêt pratique. La division fonctionnelle la plus pertinente du cervelet découle de la terminaison des afférences sur le cortex cérébelleux et de l'organisation corticonucléaire.
Division fonctionnelle (figure 1.36)
À l'image du cerveau, le cervelet comporte de la substance blanche, de la substance grise corticale et de la substance grise centrale, les trois paires de noyaux cérébelleux. Le noyau fastigial est situé dans le vermis à proximité du toit du 4e ventricule, les noyaux interposés, globuleux et emboliformes sont localisés dans la partie la plus médiale de l'hémisphère et les noyaux dentés sont situés plus latéralement, enchâssés dans la substance blanche.
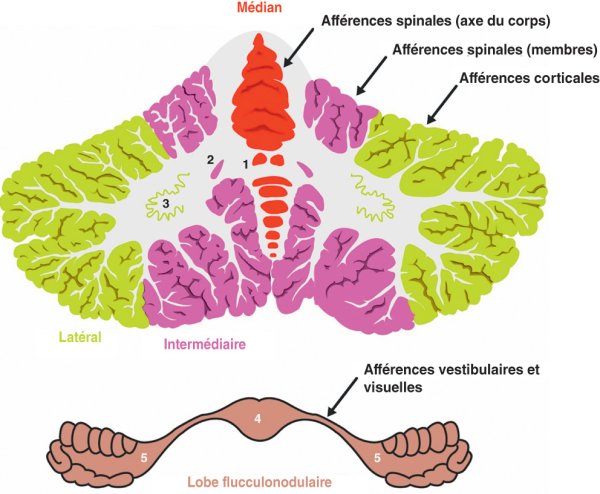
Figure 1.36
Division fonctionnelle du cervelet.
1. Noyaux fastigiaux. 2. Noyaux interposés. 3. Noyau denté. 4. Nodule. 5. Floccule.
Ainsi, dans le plan sagittal, trois zones fonctionnelles corticonucléaires peuvent être définies de manière bilatérale :
- le cervelet médian correspond au cortex du vermis associé au noyau fastigial ;
- intermédiaire associe le cortex de la partie médiale de l'hémisphère aux noyaux interposés ;
- le cervelet latéral regroupe le cortex de la partie latérale de l'hémisphère et le noyau denté.
On ajoute à ces trois grandes zones le lobe flocculonodulaire qui est formé par le cortex d'une structure impaire vermienne, le nodule, et de ses prolongements bilatéraux, les floccules.
Du fait de leurs afférences spinales prédominantes, les cervelets médian et intermédiaire sont également appelés spinocérébellum. Leur fonction principale est le contrôle du bon déroulement du mouvement en cours. Le cervelet latéral est également appelé cérébrocérébellum du fait de ses afférences corticales et régit principalement la programmation motrice. Le lobe flocculonodulaire, qui reçoit principalement des afférences vestibulaires, est défini également comme le vestibulocérébellum, et exerce son action principale sur le maintien de l'équilibre.
Contrôle de la motricité des membres (figure 1.37)
Ce contrôle est sous la dépendance du cervelet intermédiaire qui reçoit des informations proprioceptives inconscientes des membres par le tractus spinocérébelleux postérieur (anciennement faisceau de Flechsig) et par le tractus spinocérébelleux antérieur (anciennement faisceau de Gowers).
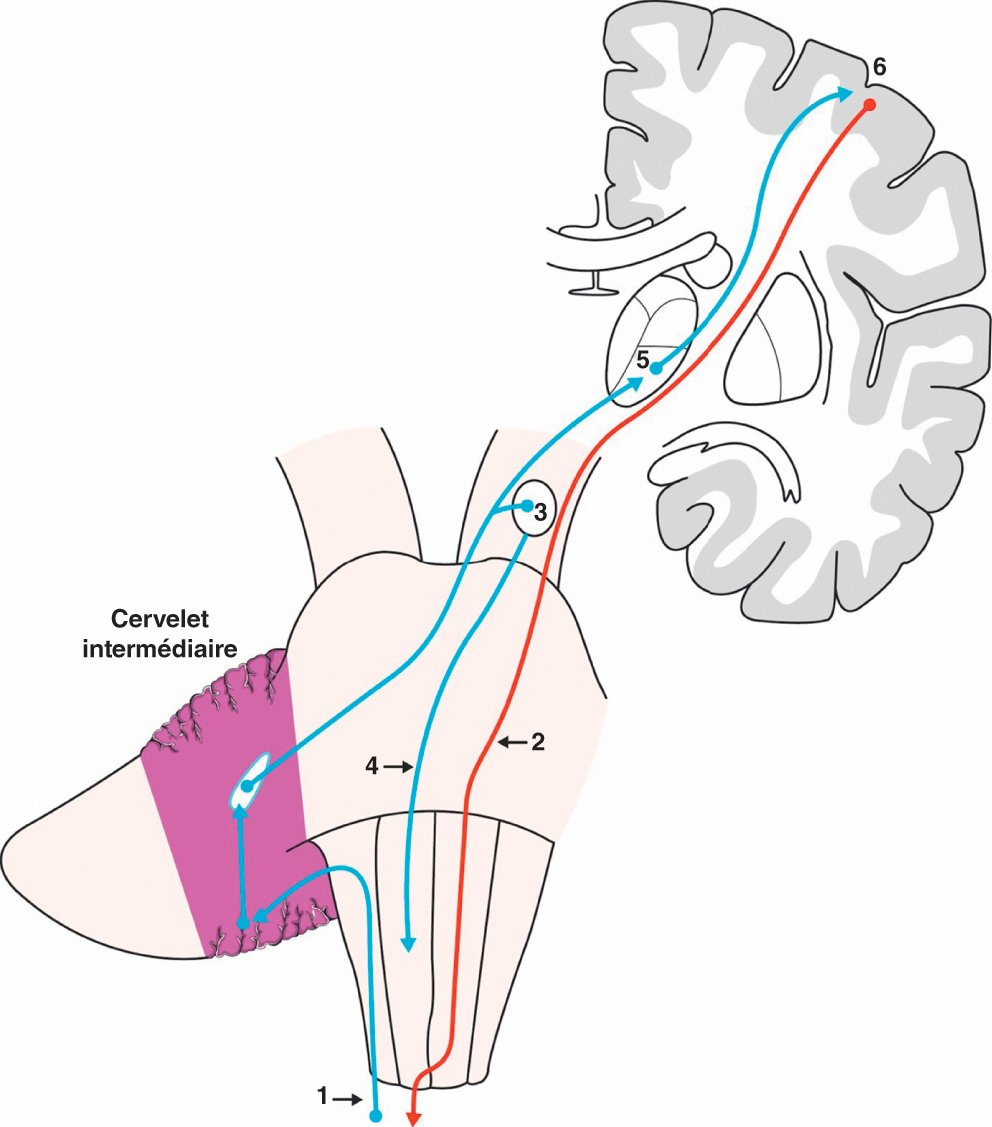
Figure 1.37
Fonctionnement du cervelet intermédiaire.
1. Afférences proprioceptives. 2. Voie motrice principale. 3. Noyau rouge. 4. Tractus rubrospinal. 5. Thalamus ventrolatéral. 6. Cortex frontal.
Depuis le cortex du cervelet intermédiaire, les fibres projettent sur les noyaux interposés puis sur les noyaux ventral antérieur et ventral latéral du thalamus. Au passage, la voie fait relais sur le noyau rouge controlatéral qui donne naissance au tractus rubrospinal qui décusse et part en direction des cornes ventrales de la moelle spinale. La voie part ensuite du thalamus en direction du cortex moteur pour effectuer sa fonction de régulation motrice. Le faisceau rubrospinal contribue également au contrôle de la motricité volontaire des membres.
Une lésion du cervelet intermédiaire aboutit à l'apparition d'un syndrome cérébelleux cinétique ipsilatéral à la lésion du fait de la double décussation des voies. L'expression clinique d'une telle lésion est l'incoordination motrice du mouvement à la fois dans ses coordonnées spatiales et temporelles (dysmétrie, asynergie, etc.). Une lésion du circuit neuronal relié au cervelet intermédiaire, dans le pont ou le thalamus par exemple, peut également aboutir à l'apparition d'un syndrome cérébelleux cinétique, ici controlatéral.
Contrôle de la motricité axiale et de l'équilibre
Les fonctions principales du cervelet latéral correspondent à la programmation du mouvement et à l'apprentissage moteur. Le cervelet latéral fonctionne en circuit fermé avec le cortex cérébral et ne reçoit pas d'afférences de la moelle spinale. Il reçoit des afférences de différentes zones corticales, en particulier des aires prémotrices et somesthésiques. Elles arrivent par les pédoncules cérébelleux moyens après un relais dans les noyaux du pont controlatéraux. Depuis le cortex du cervelet latéral, la projection se fait vers le noyau denté, puis vers les noyaux thalamiques ventral antérieur et ventral latéral avant de rejoindre le cortex cérébral. Il existe également un relais dans le noyau rouge controlatéral.
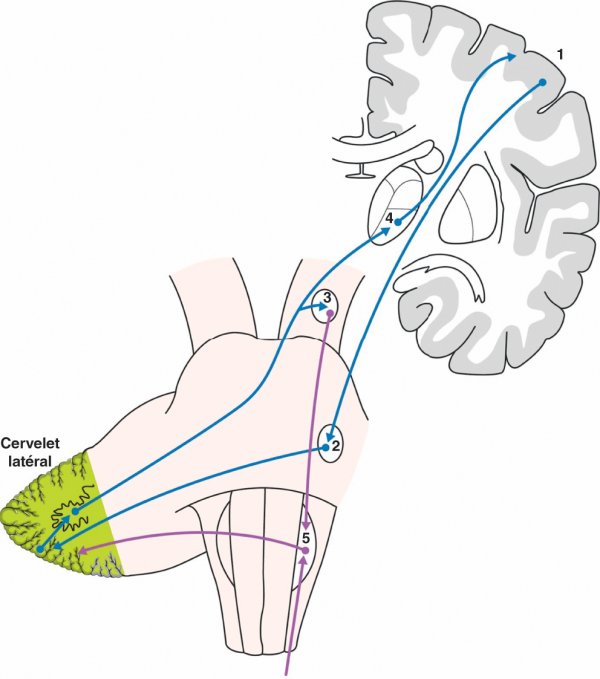
Figure 1.38
Fonctionnement du cervelet latéral.
1. Cortex frontal. 2. Noyaux du pont. 3. Noyau rouge. 4. Thalamus ventrolatéral. 5. Olive bulbaire.
Le cervelet latéral intervient très tôt dans la genèse du mouvement au moment de sa programmation et avant l'intervention du cervelet intermédiaire qui supervise l'exécution du mouvement en cours. L'échange permanent entre le cervelet latéral et les aires prémotrices permet la transition harmonieuse entre un mouvement et le mouvement suivant.
L'olive bulbaire occupe une place particulière dans le fonctionnement du cervelet latéral. Elle reçoit des afférences de la moelle spinale par le tractus spino-olivaire et du cervelet via le noyau rouge par le faisceau rubro-olivaire. L'olive bulbaire agit comme un comparateur entre le mouvement attendu et le mouvement réellement produit et envoie des informations correctrices au cervelet latéral controlatéral par le pédoncule cérébelleux inférieur. Elle intervient également dans l'apprentissage moteur.
Une lésion du cervelet latéral est généralement associée à celle du cervelet intermédiaire et aboutit à l'apparition d'un syndrome cérébelleux cinétique ipsilatéral à la lésion du fait de la double décussation des voies. L'expression clinique d'une telle lésion aboutit également à une décomposition du mouvement et à une augmentation du délai de déclenchement ou de fin de mouvement.
Anatomie fonctionnelle des ganglions de la base
Les ganglions de la base constituent un ensemble de noyaux sous-corticaux de substance grise. Ces noyaux interconnectés fonctionnant en boucles sont principalement impliqués dans le contrôle de la motricité mais ils interviennent également dans l'apprentissage moteur, le comportement, la prise de décision et les émotions. La boucle motrice est la mieux connue. Les ganglions de la base regroupent principalement un ensemble de structures anatomiques paires : le noyau caudé, le noyau lentiforme, le nucleus accumbens, le noyau subthalamique et la substance noire mésencéphalique.
Boucles motrices
Dans les modèles de fonctionnement en boucles, les ganglions de la base sont classés en structures d'entrée, structures de sortie, structures intrinsèques de traitement de l'information et structures de modulation :
- les structures d'entrée reçoivent des informations venant principalement du cortex cérébral. Elles correspondent au striatum qui regroupe le putamen, le noyau caudé et le nucleus accumbens ;
- les structures de sortie envoient des informations vers leurs cibles principalement le thalamus ventral et le tronc cérébral. Elles correspondent au globus pallidus interne et à la substance noire réticulaire ;
- les structures intrinsèques relaient et traitent les informations entre les structures d'entrée et de sortie. Elles englobent le globus pallidus externe et le noyau subthalamique ;
- une structure de modulation, la substance noire compacte, ajuste le fonctionnement des circuits internes des ganglions de la base par la voie appelée nigrostriée allant de la substance noire au striatum.
La boucle cortico-sous-corticale motrice prend naissance dans les aires prémotrices, motrice supplémentaire et dorsolatérales du lobe frontal et projette sur le striatum avec une action stimulatrice médiée par le neurotransmetteur glutamate. Après traitement des informations par les ganglions de la base, le retour se fait par relais dans les noyaux thalamiques ventral antérieur et ventral latéral vers les aires corticales de départ. Par son action stimulatrice sur les aires associatives frontales, cette boucle motrice influence directement la planification du mouvement, et le réglage des schémas moteurs complexes et indirectement le cortex moteur primaire, point de départ des voies corticospinale et corticobulbaire. Elle agit à la fois sur les mouvements volontaires des membres et sur le tonus et la posture des muscles axiaux.
Voies directe et indirecte
Le modèle de traitement de l'information par les structures intermédiaires repose sur l'existence de deux voies de traitement de l'information, la voie directe et la voie indirecte, placées entre les structures d'entrée (striatum) et de sortie (globus pallidus interne et substance noire réticulaire) des ganglions de la base.
La voie directe est représentée par les projections directes monosynaptiques, inhibitrices, gabaergiques, du striatum sur les structures de sortie. Finalement, le striatum renforce l'inhibition au niveau du globus pallidus interne et de la substance noire réticulaire induisant une facilitation dans leurs structures cibles : le thalamus, le colliculus supérieur et le tronc cérébral. Cette voie facilite donc globalement le mouvement. Par l'intermédiaire des neurones gabaergiques de la voie directe qui expriment des récepteurs dopaminergiques D1, la dopamine permet une régulation fine par un effet facilitateur sur la voie directe (figure 1.39).
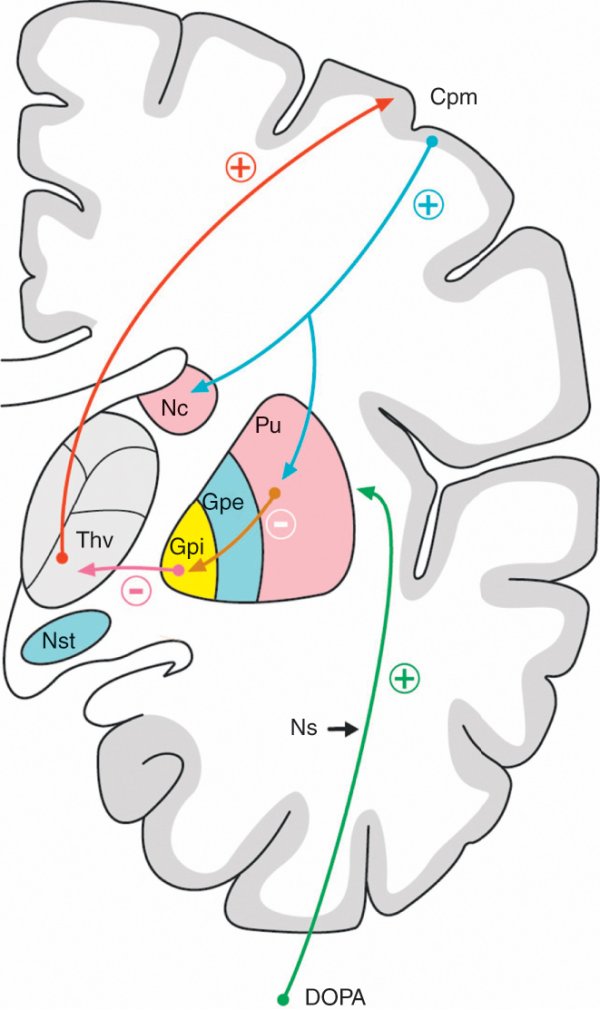
Figure 1.39
Voie directe des ganglions de la base.
Cpm : cortex prémoteur ; Gpe : globus pallidus externe ; Gpi : globus pallidus interne ; Nc : noyau caudé ; Ns : voie nigrostriée ; Nst : noyau subthalamique ; Pu : putamen ; Thv : thalamus ventral.
La voie indirecte comporte une étape supplémentaire passant par le noyau subthalamique. Le striatum émet des projections gabaergiques inhibitrices vers le globus pallidus externe d'où partent des projections gabaergiques inhibitrices vers le noyau subthalamique. Cette double inhibition stimule donc les neurones glutamatergiques excitateurs du noyau subthalamique sur les structures de sortie. Par cette voie, le striatum renforce finalement l'inhibition des structures de sortie des ganglions de la base sur leurs cibles. La voie indirecte est donc frénatrice du mouvement. La dopamine produit une inhibition sur la voie indirecte par les neurones gabaergiques qui expriment des récepteurs dopaminergiques D2 (figure 1.40).
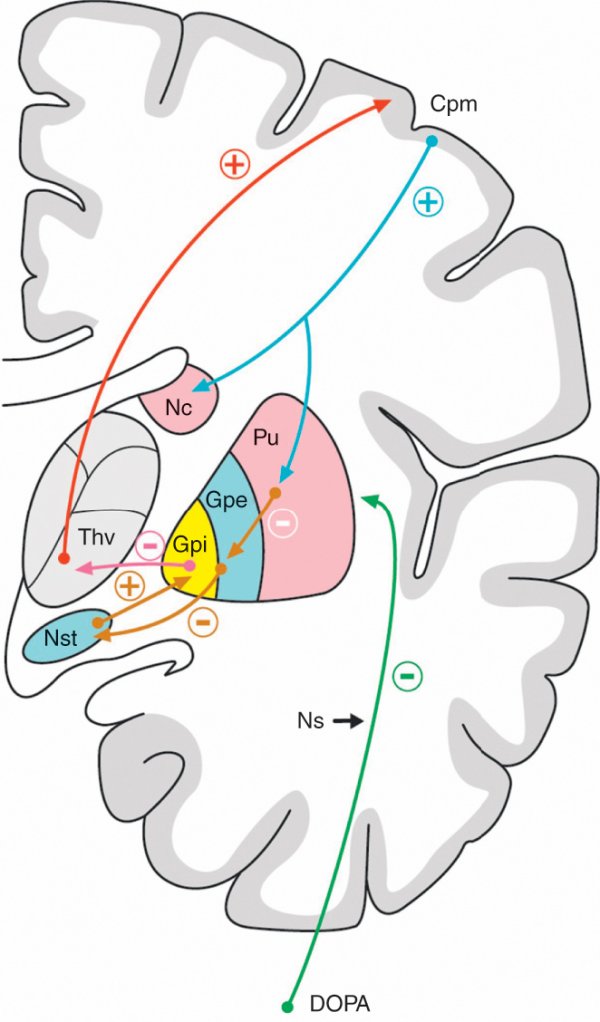
Figure 1.40
Voie indirecte des ganglions de la base.
Cpm : cortex prémoteur ; Gpe : globus pallidus externe ; Gpi : globus pallidus interne ; Nc : noyau caudé ; Ns : voie nigrostriée ; Nst : noyau subthalamique ; Pu : putamen ; Thv : thalamus ventral.
Un fonctionnement moteur adapté nécessite un juste équilibre entre la voie directe productrice de mouvements et la voie indirecte inhibitrice. Une prépondérance d'une des deux voies aboutit à l'apparition d'une pathologie de type maladie de Parkinson (excès d'inhibition) ou chorée de Huntington (excès de stimulation).